7.1 — Organização geral do sistema de endomembranas #
A emergência das células eucariontes representou um salto qualitativo na organização da vida, marcado pela internalização de funções e pela compartimentalização de processos bioquímicos que, até então, ocorriam de forma difusa no citoplasma ancestral. Essa reorganização permitiu ganhos significativos de eficiência, controle e especialização funcional, estabelecendo uma arquitetura celular na qual diferentes reações ocorrem em ambientes fisicamente delimitados e quimicamente ajustados. No entanto, essa mesma compartimentalização impõe um desafio estrutural e funcional fundamental: como integrar compartimentos distintos em um sistema coerente, dinâmico e funcionalmente interdependente. A resposta a esse desafio reside no sistema de endomembranas, um conjunto integrado de organelas e estruturas membranosas que operam como uma rede contínua de síntese, modificação, transporte e degradação de moléculas dentro da célula.
O sistema de endomembranas não deve ser compreendido como um conjunto estático de compartimentos isolados, mas como uma rede dinâmica e altamente coordenada, na qual membranas, proteínas e lipídios circulam continuamente por meio de processos de brotamento, transporte e fusão vesicular. Essa rede inclui o retículo endoplasmático, o complexo de Golgi, endossomos, lisossomos e vesículas de transporte, além de sua interface funcional com a membrana plasmática. Embora essas estruturas apresentem identidades bioquímicas e funcionais distintas, todas compartilham uma origem comum e mantêm interconexões funcionais que garantem a continuidade do fluxo intracelular de materiais [Figura].
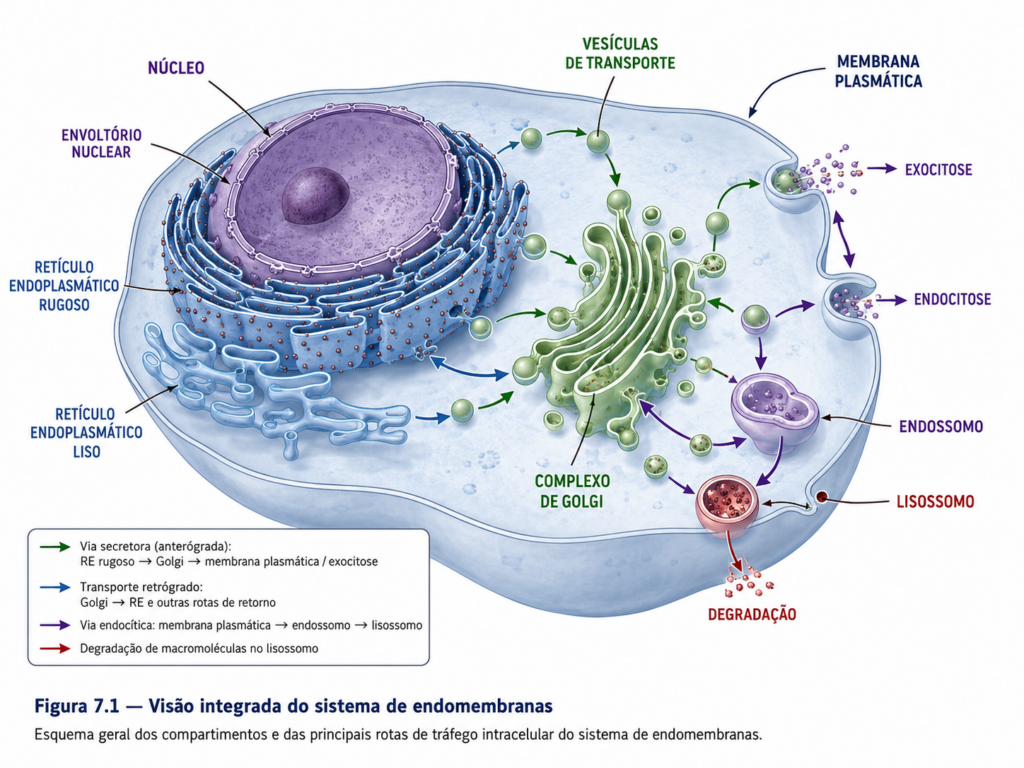
Do ponto de vista estrutural, o retículo endoplasmático representa o principal ponto de entrada de proteínas destinadas à via secretora, sendo contínuo com a membrana nuclear e funcionando como uma plataforma inicial para a síntese e o processamento de proteínas e lipídios. A partir desse compartimento, moléculas são direcionadas ao complexo de Golgi, onde sofrem modificações adicionais e são classificadas conforme seu destino final. Esse fluxo não ocorre por difusão simples, mas por meio de vesículas delimitadas por membrana, que garantem especificidade e direcionalidade ao transporte intracelular. Esse princípio de transporte vesicular estabelece a base operacional do sistema de endomembranas, permitindo que a célula mantenha compartimentos distintos sem comprometer a integração funcional.
A identidade de cada compartimento do sistema de endomembranas é definida por um conjunto específico de proteínas residentes, lipídios de membrana e condições físico-químicas, como pH e potencial redox. Essas características determinam não apenas a função de cada organela, mas também a direção e o destino das moléculas transportadas. Por exemplo, o ambiente oxidante do retículo endoplasmático favorece a formação de pontes dissulfeto em proteínas, enquanto o ambiente ácido dos lisossomos é essencial para a atividade de enzimas hidrolíticas. Essa especialização funcional evidencia que o sistema de endomembranas opera como um gradiente organizado de compartimentos, no qual moléculas são progressivamente modificadas ao longo de sua trajetória intracelular [Tabela].
A dinâmica do sistema depende criticamente da capacidade de formar e direcionar vesículas de transporte. Essas vesículas são revestidas por proteínas específicas que reconhecem e selecionam cargas moleculares, deformam a membrana para gerar brotamento e garantem o direcionamento correto até o compartimento alvo. Uma vez alcançado o destino, mecanismos de reconhecimento molecular promovem a fusão da vesícula com a membrana do compartimento receptor, permitindo a entrega do conteúdo transportado. Esse ciclo contínuo de brotamento e fusão não apenas mantém o fluxo intracelular, mas também contribui para a renovação e a manutenção das próprias membranas dos compartimentos.
A integração do sistema de endomembranas com a membrana plasmática amplia sua importância funcional, estendendo sua atuação para além do interior celular. Processos como exocitose e endocitose estabelecem uma interface direta entre o meio intracelular e o ambiente externo, permitindo a secreção de moléculas, a internalização de partículas e a regulação da composição da membrana celular. Dessa forma, o sistema de endomembranas não apenas organiza o espaço intracelular, mas também media a interação da célula com seu ambiente, desempenhando papel central na comunicação, na defesa e na adaptação celular.
Sob uma perspectiva funcional integrada, o sistema de endomembranas pode ser interpretado como uma infraestrutura logística celular, responsável por coordenar a produção, o processamento, o direcionamento e a degradação de biomoléculas. Essa visão destaca a célula como um sistema operacional complexo, no qual a eficiência depende da precisão e da sincronização dos fluxos intracelulares. Alterações nesse sistema comprometem diretamente a homeostase celular e estão associadas a diversas disfunções fisiológicas, evidenciando sua relevância para a manutenção da vida em nível celular [Quadro].
A compreensão do sistema de endomembranas, portanto, não se limita à descrição de suas estruturas, mas exige a integração de conceitos de organização espacial, dinâmica de membranas e controle molecular. Esse entendimento estabelece a base para a análise dos processos específicos que serão detalhados nas seções subsequentes, nas quais o fluxo intracelular será explorado em seus níveis estruturais, moleculares e funcionais, consolidando a visão da célula eucarionte como um sistema altamente organizado e dinamicamente integrado.
7.2 — Retículo endoplasmático: estrutura e especialização funcional #
A internalização das funções celulares nas células eucariontes não apenas fragmentou o espaço citoplasmático em compartimentos distintos, mas também exigiu a criação de superfícies especializadas capazes de sustentar fluxos intensos de síntese e processamento molecular. Nesse contexto, o retículo endoplasmático emerge como o maior e mais versátil compartimento membranoso da célula, ocupando uma fração significativa do citoplasma e estabelecendo uma interface funcional direta com o núcleo, o citosol e o restante do sistema de endomembranas. Sua organização não é aleatória, mas reflete uma arquitetura adaptativa que permite à célula integrar síntese proteica, metabolismo lipídico e controle de qualidade molecular em um único sistema contínuo.
O retículo endoplasmático é estruturalmente constituído por uma rede interconectada de túbulos e cisternas membranosas que delimitam um espaço interno denominado lúmen. Essa rede apresenta continuidade física com a membrana nuclear externa, o que estabelece uma conexão direta entre os processos de transcrição nuclear e a tradução de proteínas destinadas ao sistema de endomembranas. Essa proximidade funcional reduz a necessidade de transporte citosólico para determinadas classes de proteínas, conferindo eficiência ao fluxo de informação genética até a síntese proteica [Figura].
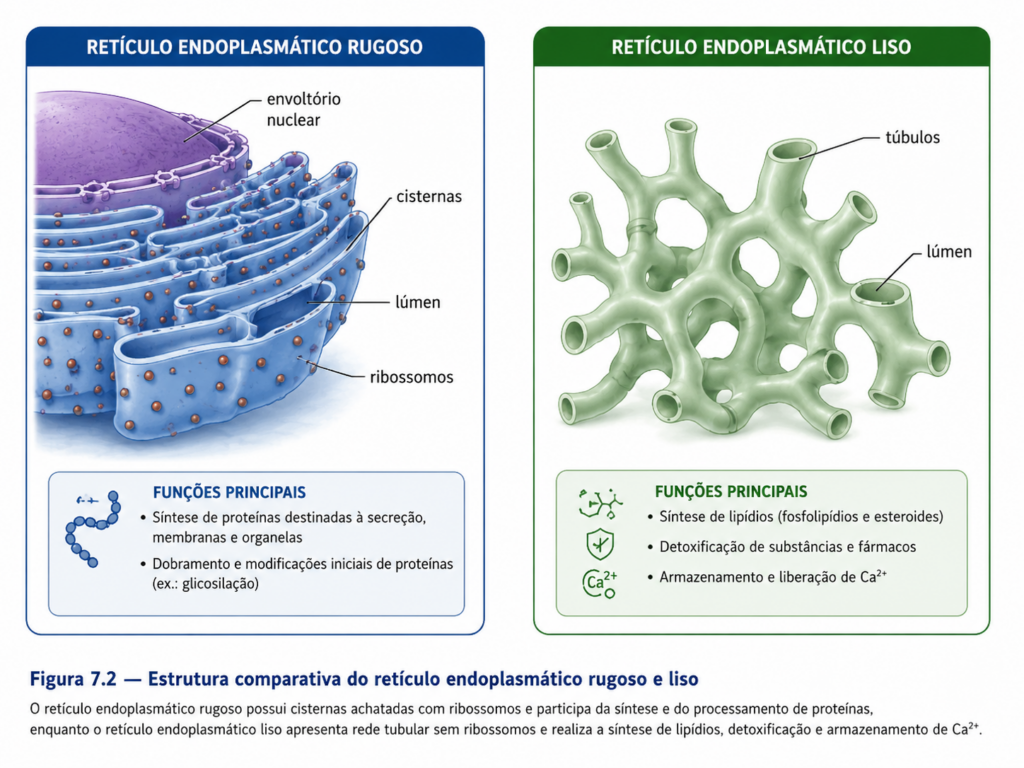
Do ponto de vista funcional, o retículo endoplasmático apresenta duas regiões morfologicamente e bioquimicamente distintas: o retículo endoplasmático rugoso (RER) e o retículo endoplasmático liso (REL). Essa distinção não representa compartimentos isolados, mas estados funcionais de uma mesma rede membranosa, cuja organização varia conforme as demandas metabólicas da célula.
O retículo endoplasmático rugoso é caracterizado pela presença de ribossomos aderidos à face citosólica de sua membrana, conferindo-lhe um aspecto granular ao microscópio eletrônico. Esses ribossomos são responsáveis pela síntese de proteínas que serão inseridas no sistema de endomembranas, destinadas à secreção, à membrana plasmática ou a compartimentos internos, como lisossomos. A associação entre ribossomos e membrana não é permanente, mas regulada por sinais moleculares presentes nas proteínas em síntese, o que permite à célula direcionar seletivamente quais polipeptídeos devem ser processados nesse compartimento.
Essa organização estabelece o RER como um ambiente especializado para a síntese co-traducional de proteínas, no qual a cadeia polipeptídica em formação é simultaneamente inserida no lúmen ou na membrana do retículo. Esse mecanismo reduz o risco de agregação proteica no citosol e permite que modificações iniciais ocorram de maneira coordenada com a síntese, estabelecendo um primeiro nível de controle sobre a qualidade estrutural das proteínas produzidas [Quadro].
Em contraste, o retículo endoplasmático liso apresenta uma morfologia predominantemente tubular e ausência de ribossomos aderidos, estando associado a funções metabólicas distintas. Entre suas principais atividades destacam-se a síntese de lipídios de membrana, incluindo fosfolipídios e esteroides, a detoxificação de compostos xenobióticos e o armazenamento e liberação de íons cálcio. Essa última função é particularmente relevante em células musculares, nas quais o retículo endoplasmático liso especializado, denominado retículo sarcoplasmático, desempenha papel central na regulação da contração celular.
A especialização funcional do REL evidencia a capacidade do retículo endoplasmático de adaptar sua estrutura às demandas fisiológicas específicas de diferentes tipos celulares. Em hepatócitos, por exemplo, observa-se uma expansão significativa do REL associada à intensa atividade de detoxificação, enquanto em células secretoras predomina o desenvolvimento do RER, refletindo a elevada produção de proteínas exportadas. Essa plasticidade estrutural demonstra que o retículo endoplasmático não é um compartimento fixo, mas um sistema dinâmico que responde continuamente às condições metabólicas e ambientais.
Além de suas funções de síntese, o retículo endoplasmático desempenha papel crítico na manutenção da homeostase celular por meio de mecanismos de controle de qualidade. Proteínas que não atingem sua conformação correta são retidas no lúmen e direcionadas para degradação, evitando o acúmulo de estruturas defeituosas que poderiam comprometer a função celular. Esse sistema de vigilância molecular é essencial para a integridade do proteoma celular e está diretamente relacionado à capacidade da célula de lidar com estresses fisiológicos e ambientais [Tabela].
A integração entre RER e REL também contribui para a expansão e renovação das membranas celulares. Lipídios sintetizados no REL são incorporados à própria membrana do retículo e posteriormente distribuídos para outros compartimentos por meio de vesículas ou transferência direta. Esse fluxo contínuo sustenta a biogênese de organelas e a manutenção da membrana plasmática, consolidando o retículo endoplasmático como o principal centro de produção de componentes estruturais da célula.
Sob uma perspectiva sistêmica, o retículo endoplasmático pode ser interpretado como o núcleo operacional do sistema de endomembranas, no qual se inicia a maior parte do fluxo intracelular de proteínas e lipídios. Sua posição estratégica, associada à sua capacidade de integrar síntese, processamento e controle de qualidade, o torna essencial para a funcionalidade global da célula eucarionte. Alterações em sua estrutura ou em seus mecanismos regulatórios repercutem diretamente sobre o funcionamento do sistema de endomembranas como um todo, evidenciando sua centralidade na organização celular.
A compreensão da estrutura e da especialização funcional do retículo endoplasmático estabelece, portanto, a base conceitual para o entendimento dos processos subsequentes de processamento, classificação e transporte de moléculas, que serão aprofundados nas próximas seções.
7.3 — Síntese, dobramento e controle de qualidade de proteínas #
A produção de proteínas destinadas ao sistema de endomembranas representa um dos processos mais críticos e energeticamente exigentes da célula eucarionte, exigindo não apenas precisão na tradução da informação genética, mas também um controle rigoroso sobre a conformação estrutural e a funcionalidade das moléculas produzidas. A simples síntese de uma cadeia polipeptídica não garante sua atividade biológica, uma vez que a função proteica depende diretamente de sua organização tridimensional. Nesse cenário, o retículo endoplasmático rugoso assume papel central ao integrar síntese, dobramento e triagem molecular em um ambiente altamente especializado, no qual a fidelidade estrutural das proteínas é continuamente monitorada.
A síntese de proteínas destinadas à via secretora inicia-se no citosol, onde ribossomos livres começam a traduzir o RNA mensageiro. No entanto, proteínas que contêm sequências sinal específicas são rapidamente reconhecidas por complexos moleculares que interrompem temporariamente a tradução e direcionam o ribossomo para a membrana do retículo endoplasmático. Esse mecanismo de direcionamento garante que a tradução prossiga de forma acoplada à inserção da cadeia polipeptídica no lúmen do retículo ou na própria membrana, estabelecendo um processo denominado síntese co-traducional [Figura].
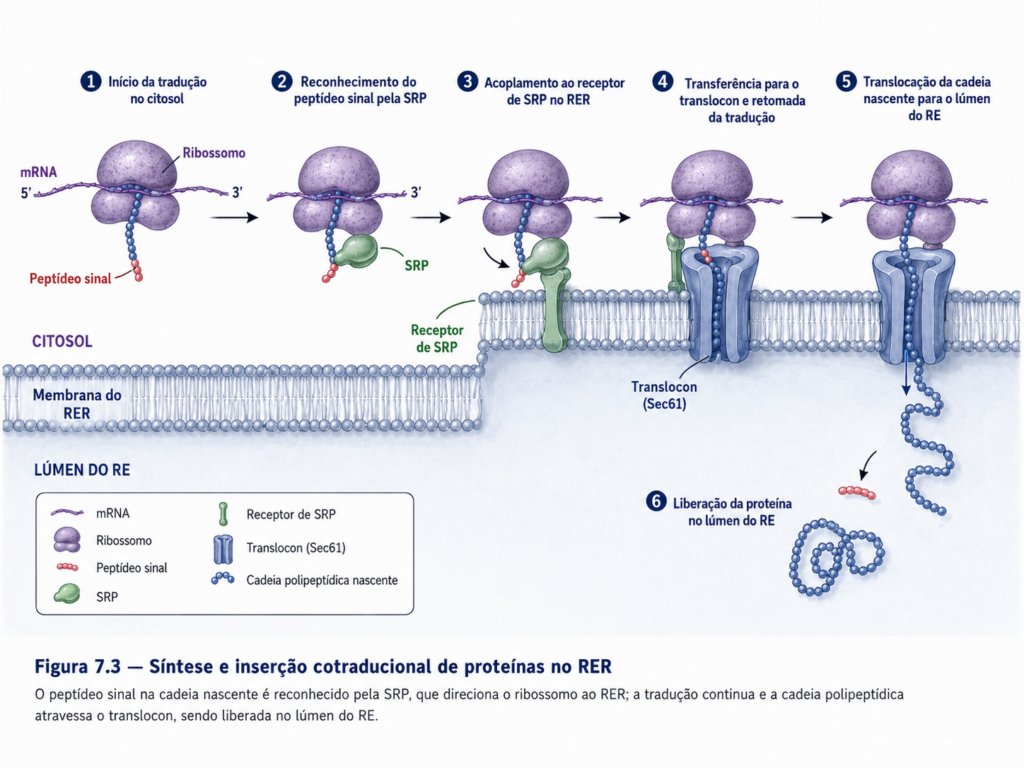
Esse acoplamento entre tradução e translocação representa um ponto crítico de controle, pois permite que a proteína em formação seja imediatamente exposta a um ambiente favorável ao dobramento correto. O lúmen do retículo endoplasmático apresenta condições específicas, incluindo concentração adequada de íons, ambiente oxidante e presença de proteínas auxiliares, que favorecem a aquisição da conformação nativa. Entre essas proteínas destacam-se as chaperonas moleculares, que interagem com polipeptídeos recém-sintetizados, prevenindo interações inadequadas e facilitando o processo de dobramento progressivo.
O dobramento proteico não é um evento espontâneo e inevitavelmente eficiente; ao contrário, trata-se de um processo sujeito a erros, especialmente em proteínas complexas ou sintetizadas em grande escala. Para mitigar esse risco, o retículo endoplasmático dispõe de um sistema altamente refinado de controle de qualidade, capaz de distinguir entre proteínas corretamente dobradas e aquelas que apresentam defeitos estruturais. Esse sistema baseia-se em ciclos de reconhecimento, modificação e reavaliação molecular, nos quais proteínas parcialmente dobradas podem ser retidas e submetidas a novas tentativas de conformação adequada [Tabela].
Entre as modificações iniciais mais relevantes que ocorrem no retículo endoplasmático destaca-se a glicosilação, processo pelo qual cadeias de oligossacarídeos são adicionadas a resíduos específicos das proteínas. Essa modificação não apenas contribui para a estabilidade estrutural, mas também atua como marcador molecular no sistema de controle de qualidade, permitindo que a célula monitore o estado de dobramento das proteínas. Proteínas que não atingem a conformação correta mantêm padrões específicos de glicosilação que as direcionam para retenção e eventual degradação.
Quando o dobramento correto não é alcançado após múltiplas tentativas, as proteínas defeituosas são direcionadas para um processo de degradação associado ao retículo endoplasmático. Nesse mecanismo, essas proteínas são retrotranslocadas para o citosol, onde são ubiquitinadas e degradadas por proteassomos. Esse sistema, conhecido como degradação associada ao retículo, desempenha papel essencial na prevenção do acúmulo de proteínas mal dobradas, que poderiam formar agregados tóxicos e comprometer a viabilidade celular [Quadro].
A eficiência desse sistema de controle é tão crítica que sua sobrecarga desencadeia respostas celulares específicas, como a resposta ao estresse do retículo endoplasmático. Quando há acúmulo excessivo de proteínas mal dobradas, a célula ativa vias de sinalização que reduzem a taxa de síntese proteica, aumentam a produção de chaperonas e intensificam os mecanismos de degradação. Esse ajuste dinâmico evidencia que o retículo endoplasmático não é apenas um local passivo de processamento, mas um centro regulador capaz de responder a variações nas condições celulares.
Além do controle de qualidade, o retículo endoplasmático também estabelece critérios de seleção para o envio de proteínas ao complexo de Golgi. Apenas proteínas corretamente dobradas e adequadamente modificadas são incorporadas em vesículas de transporte, garantindo que o fluxo intracelular seja composto por moléculas funcionalmente competentes. Esse processo assegura a integridade do sistema de endomembranas como um todo, evitando a propagação de defeitos estruturais ao longo da via secretora.
Sob uma perspectiva integrada, a síntese, o dobramento e o controle de qualidade de proteínas no retículo endoplasmático constituem um sistema altamente coordenado que combina precisão molecular com mecanismos robustos de vigilância. Esse sistema garante que a informação genética seja traduzida em estruturas proteicas funcionais, preservando a homeostase celular e sustentando processos essenciais como secreção, comunicação celular e renovação de membranas. Qualquer falha nesse equilíbrio repercute amplamente na fisiologia celular, reforçando o papel central do retículo endoplasmático na organização funcional da célula eucarionte.
7.4 — Complexo de Golgi: processamento e direcionamento molecular #
A síntese proteica no retículo endoplasmático estabelece apenas a etapa inicial de um fluxo intracelular mais amplo, no qual as moléculas recém-produzidas ainda carecem de modificações estruturais adicionais e, sobretudo, de um direcionamento preciso para seus destinos finais. A existência de múltiplos compartimentos celulares com funções distintas impõe à célula o desafio de identificar, classificar e encaminhar corretamente cada proteína ou lipídio sintetizado. Nesse contexto, o complexo de Golgi emerge como o principal centro de processamento e triagem molecular do sistema de endomembranas, operando como uma interface estratégica entre a produção inicial e a distribuição funcional dos componentes celulares.
O complexo de Golgi é estruturalmente organizado em pilhas de cisternas membranosas achatadas, dispostas de forma polarizada, o que reflete sua dinâmica funcional. Essa polaridade é definida por três regiões principais: a face cis, voltada para o retículo endoplasmático; as cisternas mediais, onde ocorrem modificações intermediárias; e a face trans, responsável pela classificação e envio das moléculas para seus destinos específicos. Essa organização não é apenas morfológica, mas funcionalmente determinante, uma vez que diferentes conjuntos de enzimas estão distribuídos ao longo dessas regiões, criando um gradiente sequencial de processamento [Figura].
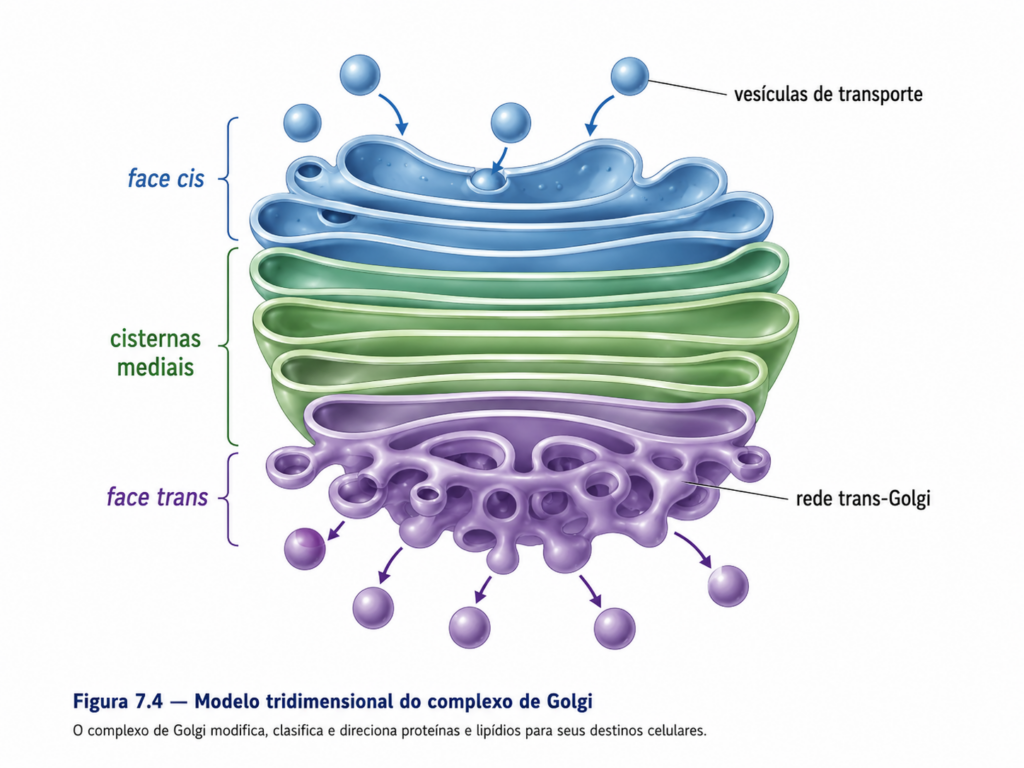
As proteínas e lipídios provenientes do retículo endoplasmático são incorporados em vesículas de transporte que se fundem com a face cis do complexo de Golgi. A partir desse ponto, essas moléculas transitam progressivamente pelas cisternas, sendo submetidas a modificações pós-traducionais que incluem principalmente a remodelação de cadeias de oligossacarídeos previamente adicionadas no retículo. Esse processo de glicosilação progressiva não ocorre de maneira uniforme, mas segue padrões específicos que variam conforme o tipo de proteína e sua função final, resultando em uma diversidade estrutural que influencia diretamente a estabilidade, a atividade e o reconhecimento molecular dessas proteínas [Tabela].
Além da glicosilação, o complexo de Golgi participa de outras modificações críticas, como a sulfatação de resíduos específicos e a fosforilação de grupos açucarados em proteínas destinadas a lisossomos. Um exemplo particularmente relevante é a adição de manose-6-fosfato em hidrolases lisossômicas, que atua como um sinal de endereçamento, permitindo que essas enzimas sejam reconhecidas por receptores específicos e direcionadas corretamente ao compartimento lisossomal. Esse mecanismo evidencia que o processamento no Golgi não se limita à modificação estrutural, mas está intrinsecamente associado à definição do destino intracelular das moléculas.
A função de triagem do complexo de Golgi atinge seu ápice na região trans, onde ocorre a separação e o empacotamento das proteínas em vesículas distintas, de acordo com seus destinos. Essas vesículas podem ser direcionadas para diferentes compartimentos, incluindo a membrana plasmática, os lisossomos ou outras regiões do sistema de endomembranas. Esse processo depende do reconhecimento de sinais moleculares presentes nas proteínas, que são interpretados por receptores específicos e mecanismos de seleção de carga. A precisão desse sistema é essencial para a manutenção da organização celular, uma vez que erros de direcionamento podem comprometer funções críticas e levar a disfunções fisiológicas [Quadro].
A dinâmica do complexo de Golgi também envolve um equilíbrio contínuo entre o fluxo anterógrado, que transporta moléculas do retículo para a periferia celular, e o fluxo retrógrado, responsável pelo retorno de proteínas residentes e componentes de membrana ao retículo ou a cisternas anteriores. Esse fluxo bidirecional é fundamental para a manutenção da identidade funcional das cisternas do Golgi, garantindo que as enzimas responsáveis pelas modificações permaneçam corretamente distribuídas ao longo do sistema. Modelos como o de maturação cisternal sugerem que as próprias cisternas evoluem ao longo do tempo, enquanto enzimas são recicladas, evidenciando a natureza altamente dinâmica desse compartimento.
A relevância funcional do complexo de Golgi se estende além do processamento intracelular, influenciando diretamente a interação da célula com seu ambiente. A composição de glicoproteínas e glicolipídios da membrana plasmática, por exemplo, depende das modificações realizadas no Golgi, afetando processos como reconhecimento celular, adesão e resposta imunológica. Em organismos multicelulares, essas modificações são essenciais para a comunicação entre células e para a organização de tecidos, demonstrando que a atividade do Golgi possui implicações que transcendem o nível celular individual.
Do ponto de vista fisiopatológico, alterações na estrutura ou na função do complexo de Golgi estão associadas a diversas doenças, muitas das quais relacionadas a defeitos na glicosilação ou no tráfego de proteínas. Essas condições evidenciam que a precisão no processamento e no direcionamento molecular é indispensável para a homeostase celular e para o funcionamento adequado de sistemas biológicos mais complexos.
Em síntese, o complexo de Golgi atua como um centro integrador no sistema de endomembranas, no qual moléculas são modificadas, classificadas e distribuídas com alta especificidade. Sua organização polarizada e sua dinâmica funcional garantem que o fluxo intracelular de proteínas e lipídios ocorra de forma ordenada e eficiente, consolidando a célula eucarionte como um sistema altamente coordenado. A compreensão desses processos estabelece a base para o entendimento dos mecanismos de transporte vesicular que serão aprofundados na sequência, nos quais a especificidade e a direção do tráfego intracelular são operacionalizadas em nível molecular.
7.5 — Tráfego vesicular: mecanismos e especificidade #
A compartimentalização da célula eucarionte, embora essencial para a especialização funcional, impõe a necessidade de um sistema eficiente de comunicação entre organelas fisicamente separadas. A simples proximidade espacial não garante a troca de materiais, exigindo mecanismos altamente controlados que permitam o transporte direcionado de proteínas, lipídios e outras moléculas entre compartimentos distintos. Nesse cenário, o tráfego vesicular constitui o principal mecanismo de integração funcional do sistema de endomembranas, permitindo que a célula mantenha simultaneamente a compartimentalização e a continuidade operacional de seus processos internos.
O tráfego vesicular baseia-se na formação de vesículas delimitadas por membrana que brotam de um compartimento doador e se deslocam até um compartimento receptor, onde se fundem para liberar seu conteúdo. Esse processo envolve uma sequência coordenada de eventos que inclui seleção de carga, deformação da membrana, formação da vesícula, transporte intracelular e fusão com o destino final. Cada uma dessas etapas é regulada por proteínas específicas que garantem precisão e direcionalidade ao fluxo intracelular [Figura].
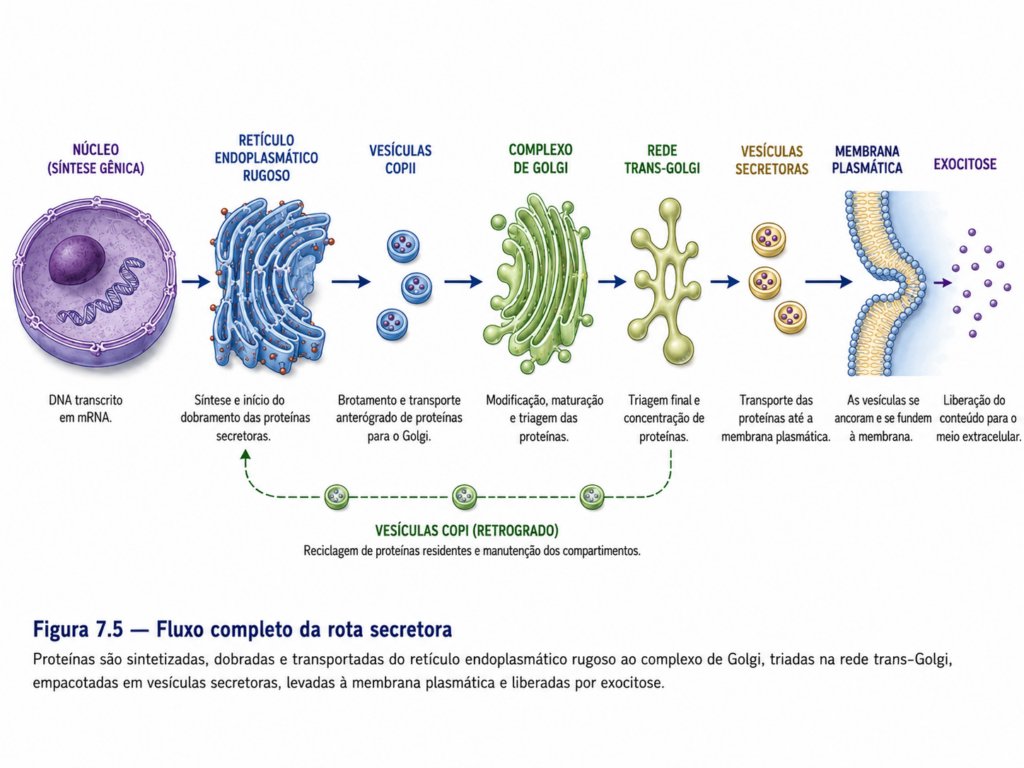
A formação de vesículas inicia-se com o recrutamento de proteínas de revestimento na membrana do compartimento doador. Essas proteínas desempenham funções estruturais e seletivas, promovendo a curvatura da membrana e a incorporação de moléculas específicas na vesícula em formação. Entre os principais sistemas de revestimento destacam-se as proteínas do tipo COPII, responsáveis pelo transporte do retículo endoplasmático para o complexo de Golgi; COPI, associadas ao transporte retrógrado dentro do Golgi e de volta ao retículo; e clatrina, envolvida em processos de endocitose e transporte a partir da face trans do Golgi. Cada um desses sistemas confere identidade funcional às vesículas e determina suas rotas intracelulares [Tabela].
A seleção de carga representa um ponto crítico de controle no tráfego vesicular. Proteínas destinadas ao transporte apresentam sinais moleculares específicos que são reconhecidos por receptores na membrana do compartimento doador. Esses receptores interagem com as proteínas de revestimento, garantindo que apenas moléculas apropriadas sejam incorporadas à vesícula. Esse mecanismo assegura que o transporte intracelular não ocorra de maneira aleatória, mas siga padrões altamente organizados que preservam a identidade funcional dos compartimentos.
Após o brotamento, as vesículas perdem seu revestimento proteico e tornam-se aptas para o transporte no citoplasma. Esse deslocamento não depende exclusivamente de difusão, sendo frequentemente mediado por proteínas motoras associadas ao citoesqueleto, como dineínas e cinesinas, que permitem o transporte direcionado ao longo de microtúbulos. Essa associação com o citoesqueleto confere eficiência e velocidade ao tráfego vesicular, especialmente em células de grande volume ou altamente polarizadas.
O reconhecimento do compartimento alvo constitui uma etapa determinante para a especificidade do tráfego vesicular. Esse processo é mediado por proteínas de ancoragem e por pequenas GTPases, como as proteínas Rab, que atuam como marcadores moleculares de identidade. Essas proteínas garantem que a vesícula seja direcionada ao compartimento correto, evitando fusões indevidas que poderiam comprometer a organização celular. Uma vez estabelecido o contato inicial, proteínas específicas promovem a aproximação das membranas e a preparação para a fusão [Quadro].
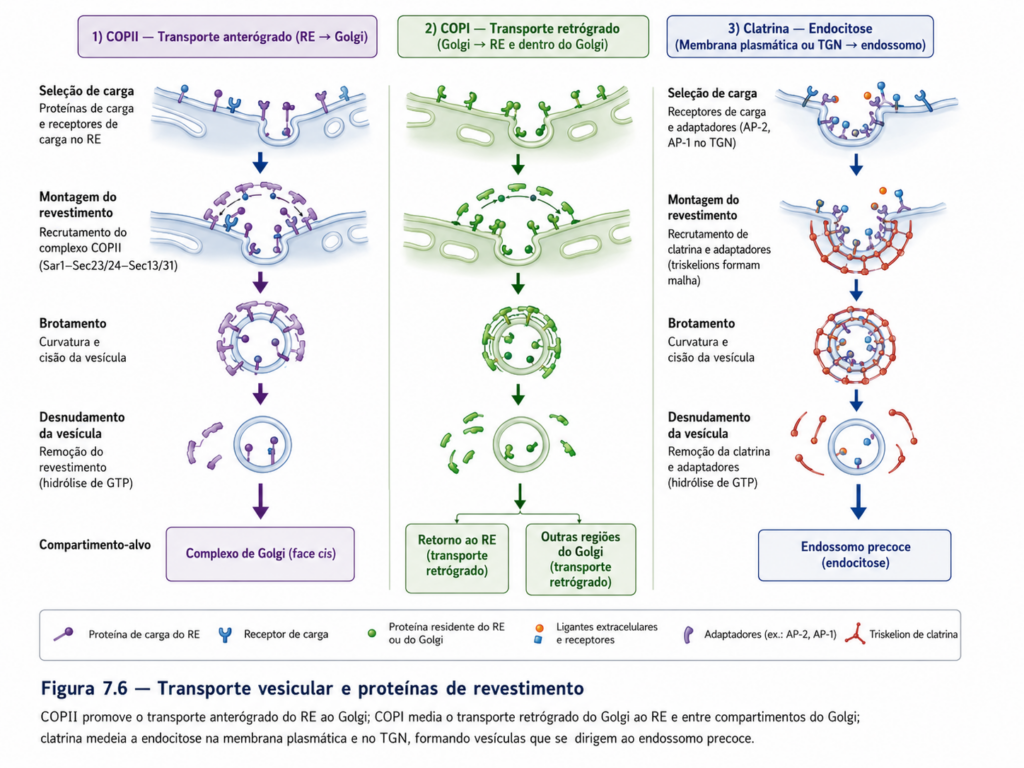
A fusão vesicular é mediada por um conjunto altamente conservado de proteínas denominadas SNAREs, que se encontram tanto na vesícula quanto na membrana alvo. A interação entre essas proteínas promove a aproximação íntima das bicamadas lipídicas, reduzindo a barreira energética necessária para a fusão e permitindo a continuidade das membranas. Esse processo culmina na liberação do conteúdo vesicular no compartimento receptor ou na incorporação de proteínas de membrana ao destino final.
A dinâmica do tráfego vesicular não se limita ao fluxo unidirecional de materiais, mas envolve um equilíbrio entre transporte anterógrado e retrógrado. Enquanto o fluxo anterógrado direciona proteínas recém-sintetizadas ao longo da via secretora, o fluxo retrógrado recupera componentes de membrana e proteínas residentes, mantendo a identidade funcional dos compartimentos. Essa bidirecionalidade é essencial para a estabilidade do sistema de endomembranas, permitindo ajustes contínuos e evitando o esgotamento de componentes críticos.
Sob uma perspectiva integrada, o tráfego vesicular pode ser compreendido como o sistema logístico da célula, no qual cada vesícula atua como uma unidade de transporte altamente especializada. A precisão desse sistema garante que proteínas e lipídios sejam entregues nos locais corretos, no momento adequado e em quantidades apropriadas, sustentando processos fundamentais como secreção, renovação de membranas, sinalização celular e degradação de biomoléculas.
A relevância desse sistema torna-se evidente quando se consideram as consequências de sua disfunção. Alterações em proteínas de revestimento, motores moleculares ou componentes do sistema de fusão podem resultar em falhas no transporte intracelular, comprometendo a homeostase celular e contribuindo para o desenvolvimento de diversas patologias. Assim, o tráfego vesicular não apenas integra o sistema de endomembranas, mas também representa um elemento central na manutenção da funcionalidade e da viabilidade celular.
7.6 — Endocitose: internalização e processamento de materiais #
A manutenção da homeostase celular não depende apenas da capacidade de sintetizar, modificar e secretar moléculas, mas também da habilidade de internalizar componentes do meio extracelular e remodelar continuamente a própria membrana plasmática. A célula eucarionte opera em um ambiente dinâmico, no qual nutrientes, sinais e partículas externas precisam ser capturados, interpretados e, em muitos casos, degradados ou reciclados. Essa necessidade estabelece a endocitose como um processo essencial de interface entre o meio externo e o sistema de endomembranas, permitindo à célula internalizar materiais de forma seletiva e integrá-los ao seu fluxo intracelular.
A endocitose compreende um conjunto de mecanismos pelos quais porções da membrana plasmática invaginam-se e formam vesículas que transportam substâncias para o interior da célula. Esse processo não é homogêneo, apresentando diferentes modalidades que variam quanto ao grau de seletividade, ao tipo de material internalizado e aos mecanismos moleculares envolvidos. Apesar dessa diversidade, todas as formas de endocitose compartilham princípios comuns relacionados à deformação da membrana, formação de vesículas e direcionamento para compartimentos intracelulares específicos [Figura].
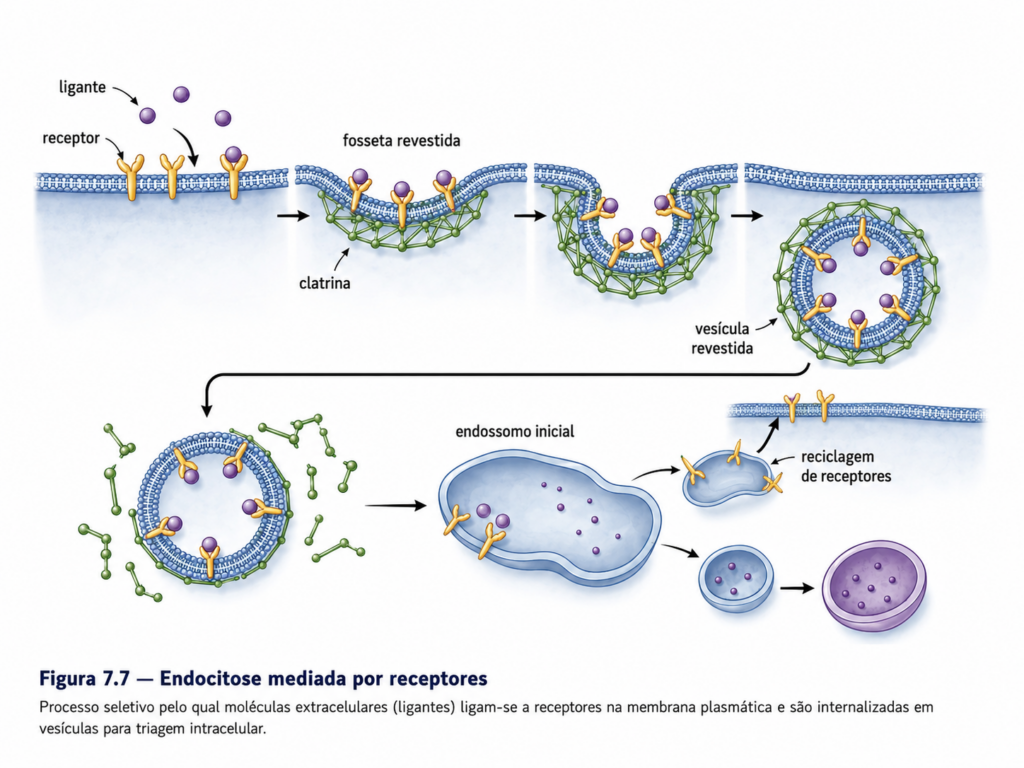
Entre os mecanismos mais refinados de endocitose destaca-se a endocitose mediada por receptores, um processo altamente seletivo que permite à célula capturar moléculas específicas presentes em baixas concentrações no meio extracelular. Nesse mecanismo, ligantes extracelulares, como hormônios, lipoproteínas ou fatores de crescimento, ligam-se a receptores específicos na superfície celular, formando complexos que se concentram em regiões especializadas da membrana. Essas regiões são revestidas por clatrina, uma proteína estrutural que promove a curvatura da membrana e a formação de vesículas revestidas, garantindo a internalização eficiente e direcionada dessas moléculas [Tabela].
Após o brotamento, as vesículas perdem seu revestimento de clatrina e se fundem com compartimentos intracelulares denominados endossomos iniciais. Esses compartimentos funcionam como centros de triagem, nos quais ocorre a separação entre receptores e ligantes. Em muitos casos, os receptores são reciclados de volta à membrana plasmática, enquanto os ligantes são direcionados para degradação ou processamento adicional. Esse mecanismo de reciclagem permite à célula manter a eficiência na captação de moléculas, evitando a necessidade de síntese contínua de novos receptores.
Além da endocitose mediada por receptores, a célula também utiliza processos menos seletivos, como a pinocitose, que envolve a internalização contínua de fluido extracelular e solutos dissolvidos. Esse mecanismo contribui para a amostragem do ambiente e para a manutenção do equilíbrio osmótico, embora não apresente o mesmo grau de especificidade observado na endocitose mediada por receptores. Em contraste, a fagocitose representa um processo especializado na internalização de partículas de grande porte, como microrganismos ou detritos celulares, sendo particularmente relevante em células do sistema imune, como macrófagos e neutrófilos.
Independentemente do mecanismo de internalização, o destino das vesículas endocíticas converge para um sistema progressivo de compartimentos endossomais, nos quais ocorre a maturação e o processamento do material internalizado. Os endossomos iniciais evoluem para endossomos tardios, caracterizados por alterações estruturais e acidificação progressiva do lúmen. Essa acidificação desempenha papel crucial na dissociação de complexos receptor-ligante e na ativação de enzimas hidrolíticas que atuarão na degradação de macromoléculas.
A etapa final desse processo ocorre nos lisossomos, compartimentos altamente especializados em digestão intracelular, nos quais enzimas hidrolíticas degradam proteínas, lipídios, carboidratos e ácidos nucleicos em seus componentes básicos. Esses produtos podem ser reutilizados pela célula em processos biossintéticos, evidenciando que a endocitose não se limita à internalização, mas integra-se a um ciclo contínuo de renovação e reaproveitamento de biomoléculas [Quadro].
A endocitose também exerce papel central na regulação da superfície celular. Ao internalizar receptores de membrana, a célula pode modular sua sensibilidade a estímulos externos, ajustando a intensidade e a duração de respostas sinalizadoras. Esse mecanismo é fundamental em processos como dessensibilização de receptores hormonais e controle de vias de sinalização, demonstrando que a endocitose participa ativamente da regulação funcional da célula.
Sob uma perspectiva integrada, a endocitose representa um componente essencial do sistema de endomembranas, estabelecendo uma via de entrada que complementa os processos de secreção e transporte intracelular. Sua capacidade de combinar seletividade, eficiência e integração funcional permite à célula interagir com seu ambiente de forma controlada, garantindo a aquisição de recursos, a remoção de partículas indesejadas e a regulação de sua própria superfície.
A compreensão dos mecanismos de endocitose revela, portanto, que a célula eucarionte não é um sistema fechado, mas uma entidade dinâmica que constantemente troca matéria e informação com o meio externo. Essa interação contínua sustenta processos fisiológicos fundamentais e estabelece a base para respostas adaptativas em contextos ambientais e patológicos, reforçando o papel central do sistema de endomembranas na organização e na funcionalidade celular.
7.7 — Exocitose e secreção celular #
A internalização de materiais por endocitose estabelece apenas metade do fluxo de troca entre a célula e seu ambiente. Em paralelo, a célula eucarionte mantém um fluxo contínuo de exportação de moléculas, essencial para a comunicação intercelular, a renovação da membrana plasmática e a manutenção de funções fisiológicas complexas. Esse processo de exportação, denominado exocitose, representa a etapa final da via secretora e integra o sistema de endomembranas ao meio extracelular, permitindo que proteínas e lipídios produzidos internamente alcancem seus destinos finais fora da célula ou na própria superfície celular.
A exocitose ocorre por meio da fusão de vesículas derivadas do complexo de Golgi com a membrana plasmática, resultando na liberação do conteúdo vesicular no espaço extracelular e na incorporação de seus componentes de membrana à superfície celular. Esse processo não apenas permite a secreção de moléculas, mas também contribui para a expansão e renovação da membrana plasmática, evidenciando sua importância estrutural além da função secretora [Figura].
Do ponto de vista funcional, a exocitose pode ser dividida em dois modos principais: secreção constitutiva e secreção regulada. A secreção constitutiva ocorre de forma contínua em praticamente todas as células, sendo responsável pela entrega constante de proteínas de membrana e componentes da matriz extracelular. Nesse mecanismo, vesículas formadas no Golgi são direcionadas diretamente à membrana plasmática sem necessidade de estímulos adicionais, garantindo a manutenção da estrutura celular e da interface com o meio externo.
Em contraste, a secreção regulada é característica de células especializadas, como células endócrinas, neurônios e células exócrinas, nas quais a liberação de moléculas ocorre de maneira controlada e dependente de sinais específicos. Nesses casos, proteínas são armazenadas em vesículas secretoras que permanecem no citoplasma até que um estímulo desencadeie sua fusão com a membrana plasmática. Esse mecanismo permite uma resposta rápida e coordenada a sinais ambientais, sendo fundamental em processos como liberação de hormônios, neurotransmissores e enzimas digestivas [Tabela].
A fusão das vesículas com a membrana plasmática é mediada por mecanismos moleculares altamente específicos, semelhantes àqueles envolvidos no tráfego vesicular intracelular. Proteínas do sistema SNARE desempenham papel central nesse processo, promovendo a aproximação e a fusão das bicamadas lipídicas. A especificidade dessas interações garante que a fusão ocorra apenas nos locais apropriados, evitando a liberação indevida de conteúdo vesicular.
Na secreção regulada, a ativação da fusão vesicular está frequentemente associada a variações na concentração intracelular de cálcio. O aumento abrupto desse íon, desencadeado por sinais extracelulares, atua como um gatilho que promove a reorganização de proteínas envolvidas na fusão, permitindo a liberação rápida do conteúdo armazenado. Esse mecanismo é particularmente evidente em sinapses neuronais, onde a entrada de cálcio desencadeia a liberação de neurotransmissores em escalas de tempo extremamente curtas [Quadro].
Além da liberação de moléculas solúveis, a exocitose também participa da inserção de proteínas e lipídios na membrana plasmática, contribuindo para a modulação de sua composição e funcionalidade. Receptores, canais iônicos e outras proteínas de membrana são continuamente adicionados ou removidos, permitindo à célula ajustar sua sensibilidade a estímulos e sua capacidade de interação com o ambiente. Essa dinâmica é essencial para processos como sinalização celular, adesão e resposta imune.
A exocitose também está diretamente associada à manutenção do equilíbrio entre entrada e saída de membranas na célula. O material incorporado por endocitose pode ser parcialmente reciclado e reinserido na membrana plasmática por meio de vias exocíticas, estabelecendo um ciclo contínuo que regula a área de superfície celular e a distribuição de componentes de membrana. Esse equilíbrio dinâmico garante que a célula mantenha sua integridade estrutural mesmo diante de intensas atividades de transporte.
Sob uma perspectiva funcional integrada, a exocitose pode ser compreendida como o mecanismo pelo qual a célula projeta suas funções para além de seus limites físicos. Ao secretar moléculas sinalizadoras, enzimas e componentes estruturais, a célula participa ativamente da organização de tecidos, da regulação fisiológica e da interação com outros organismos. Esse papel torna a exocitose um processo central não apenas para a célula individual, mas para o funcionamento de sistemas biológicos complexos.
Alterações nos mecanismos de exocitose estão associadas a diversas disfunções, incluindo distúrbios neurológicos, endocrinológicos e imunológicos, evidenciando que a precisão desse processo é fundamental para a homeostase do organismo. A compreensão da exocitose, portanto, não se limita à sua descrição mecânica, mas envolve a análise de sua integração com processos de sinalização, tráfego vesicular e organização celular.
A exocitose consolida, assim, a célula eucarionte como um sistema aberto e altamente interativo, no qual fluxos contínuos de matéria e informação conectam o interior celular ao ambiente externo. Essa conexão estabelece a base para a coordenação de processos fisiológicos em múltiplas escalas, reforçando a centralidade do sistema de endomembranas na dinâmica funcional da vida celular.
7.8 — Lisossomos e digestão intracelular #
A internalização de materiais por endocitose e a produção contínua de componentes celulares estabelecem uma demanda permanente por sistemas capazes de degradar, reciclar e reorganizar biomoléculas. A célula eucarionte não apenas sintetiza e exporta estruturas, mas também precisa remover componentes danificados, processar partículas internalizadas e reutilizar seus constituintes básicos. Nesse contexto, os lisossomos emergem como compartimentos especializados em digestão intracelular, desempenhando papel central na renovação estrutural e na manutenção da homeostase celular.
Os lisossomos são organelas delimitadas por membrana que contêm um conjunto diversificado de enzimas hidrolíticas capazes de degradar praticamente todos os tipos de macromoléculas biológicas, incluindo proteínas, lipídios, carboidratos e ácidos nucleicos. Essas enzimas apresentam atividade ótima em pH ácido, característica mantida por sistemas de bombeamento de prótons localizados na membrana lisossomal, que acidificam o lúmen do compartimento. Essa condição não apenas favorece a atividade enzimática, mas também contribui para a segurança celular, uma vez que limita a ação dessas enzimas ao interior do lisossomo [Figura].
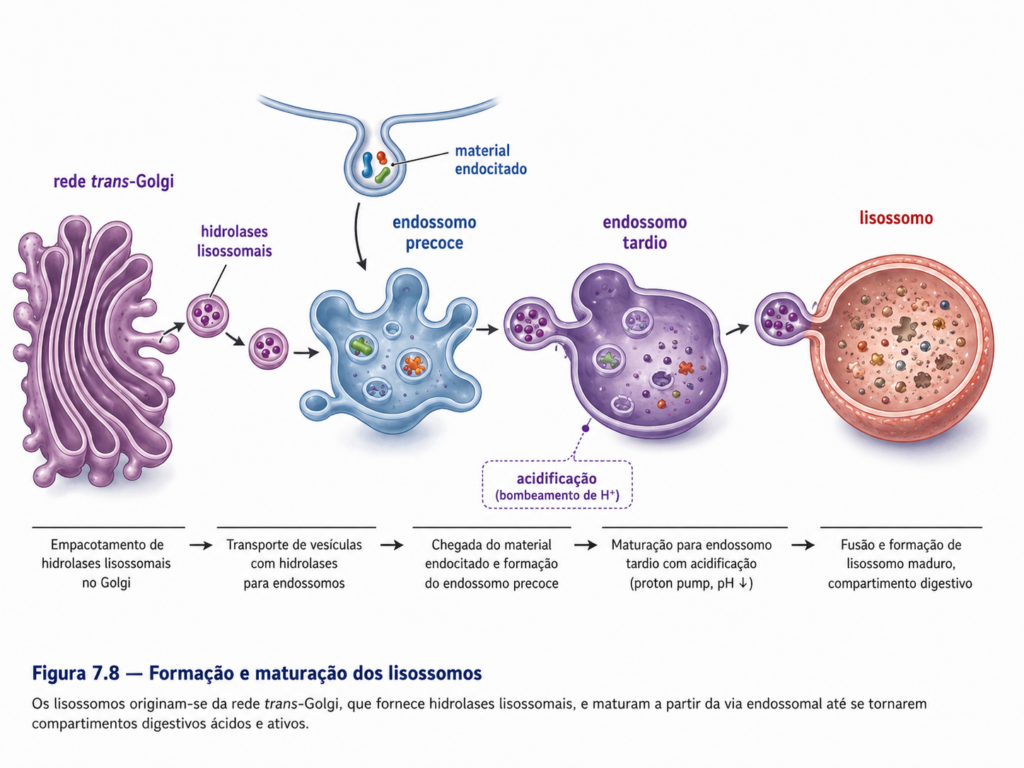
A origem dos lisossomos está diretamente associada ao sistema de endomembranas, envolvendo a contribuição do retículo endoplasmático, do complexo de Golgi e de compartimentos endossomais. Enzimas lisossomais são sintetizadas no retículo endoplasmático e posteriormente modificadas no Golgi, onde recebem sinais moleculares específicos que permitem seu direcionamento correto. Essas enzimas são então incorporadas a vesículas que se fundem com endossomos, dando origem a estruturas que evoluem funcionalmente até adquirir características lisossomais completas. Esse processo evidencia que os lisossomos não são compartimentos isolados, mas resultam de uma dinâmica contínua de fusão e maturação dentro do sistema de endomembranas [Tabela].
A digestão intracelular mediada por lisossomos pode ser compreendida a partir de três principais vias: heterofagia, autofagia e endocitose. Na heterofagia, materiais provenientes do meio extracelular são internalizados por endocitose ou fagocitose e direcionados aos lisossomos para degradação. Esse processo é essencial para a obtenção de nutrientes e para a defesa contra agentes patogênicos. Na autofagia, a célula direciona seus próprios componentes para degradação, envolvendo a formação de vesículas que englobam organelas ou regiões citoplasmáticas e posteriormente se fundem com lisossomos. Esse mecanismo desempenha papel fundamental na renovação celular e na adaptação a condições de estresse.
A capacidade de degradar estruturas internas permite à célula eliminar organelas disfuncionais, reciclar macromoléculas e ajustar sua composição estrutural conforme as demandas metabólicas. Esse processo não ocorre de maneira aleatória, mas envolve mecanismos regulatórios que reconhecem estruturas a serem degradadas e direcionam sua incorporação em vesículas autofágicas. A integração entre autofagia e atividade lisossomal evidencia que a digestão intracelular é um componente ativo da regulação celular, contribuindo para a adaptação a mudanças ambientais e para a sobrevivência em condições adversas [Quadro].
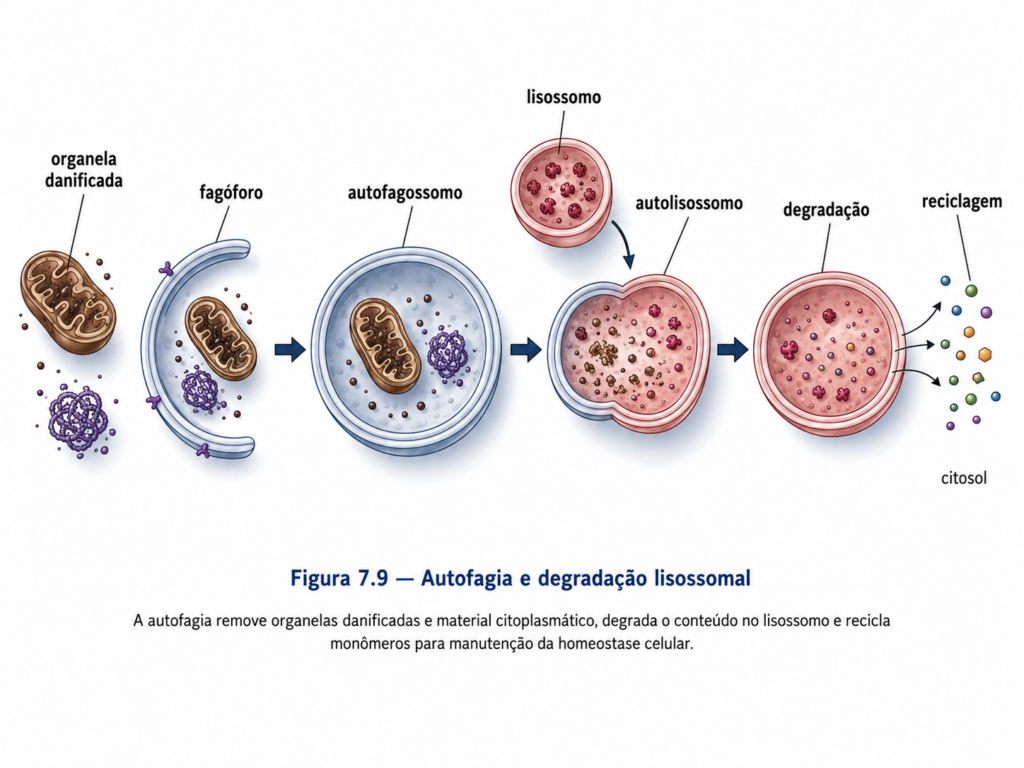
Após a degradação, os produtos resultantes, como aminoácidos, ácidos graxos e açúcares simples, são transportados de volta ao citosol, onde podem ser reutilizados em processos biossintéticos ou como fonte de energia. Esse ciclo de degradação e reaproveitamento confere eficiência metabólica à célula, reduzindo a dependência de recursos externos e permitindo a manutenção da homeostase em diferentes condições fisiológicas.
Além de sua função degradativa, os lisossomos participam de processos celulares mais amplos, incluindo regulação metabólica, sinalização intracelular e até mesmo mecanismos de morte celular. A integridade da membrana lisossomal é crucial para o funcionamento adequado da célula, uma vez que a liberação descontrolada de enzimas hidrolíticas no citosol pode desencadear danos extensivos e levar à morte celular. Esse potencial destrutivo controlado reforça a importância de mecanismos que preservem a compartimentalização e a regulação da atividade lisossomal.
Do ponto de vista fisiopatológico, disfunções nos lisossomos estão associadas a um conjunto de doenças conhecidas como doenças de armazenamento lisossomal, caracterizadas pelo acúmulo de substratos não degradados devido à deficiência de enzimas específicas. Essas condições evidenciam que a eficiência da digestão intracelular é indispensável para o funcionamento adequado da célula e do organismo como um todo.
Sob uma perspectiva integrada, os lisossomos podem ser compreendidos como centros de reciclagem celular, nos quais a matéria orgânica é continuamente transformada e reinserida no metabolismo celular. Sua atuação garante a renovação estrutural, a eliminação de componentes danificados e a adaptação a variações ambientais, consolidando o sistema de endomembranas como uma rede funcional capaz de equilibrar síntese, transporte e degradação.
A compreensão dos lisossomos e da digestão intracelular completa a visão do sistema de endomembranas como um ciclo contínuo de produção, processamento e reciclagem, no qual a célula mantém sua integridade estrutural e funcional por meio de fluxos dinâmicos e altamente regulados.
7.9 — Integração funcional do sistema de endomembranas #
A compartimentalização da célula eucarionte estabelece ambientes especializados para a realização de funções específicas, mas a eficiência desse modelo depende da existência de mecanismos capazes de integrar esses compartimentos em um sistema operacional coerente. A síntese de proteínas no retículo endoplasmático, seu processamento no complexo de Golgi, a internalização por endocitose e a degradação em lisossomos não ocorrem como eventos isolados, mas como etapas interdependentes de um fluxo contínuo de matéria e informação. Essa interdependência define o sistema de endomembranas como uma rede funcional integrada, na qual a identidade e a dinâmica de cada compartimento são mantidas por fluxos coordenados de membranas e moléculas.
A integração funcional do sistema de endomembranas depende, em primeiro lugar, da manutenção da identidade dos compartimentos celulares. Cada organela apresenta uma composição específica de proteínas de membrana, lipídios e condições físico-químicas que determinam sua função. Essa identidade não é fixa, sendo constantemente preservada por mecanismos de transporte seletivo que garantem a entrega correta de componentes e a remoção de elementos inadequados. O equilíbrio entre fluxo anterógrado e retrógrado é essencial nesse contexto, pois permite que proteínas residentes sejam recicladas e que as características funcionais de cada compartimento sejam continuamente restauradas [Figura].
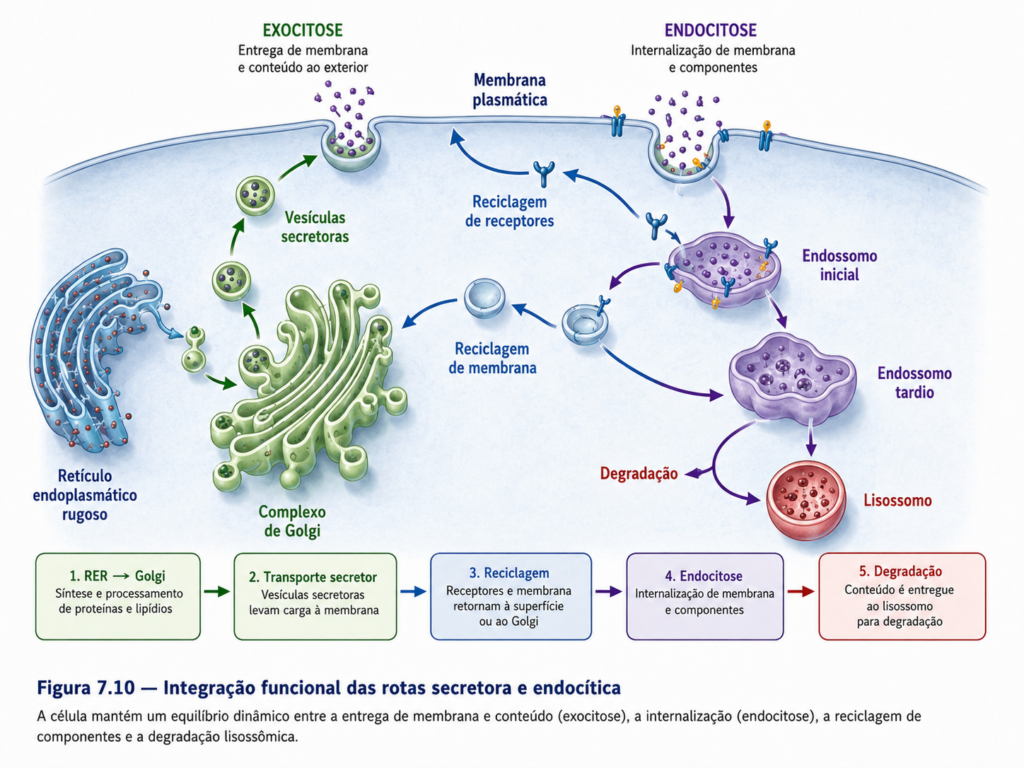
O fluxo anterógrado, que se inicia no retículo endoplasmático e segue em direção ao complexo de Golgi e à membrana plasmática, representa a via principal de exportação de proteínas e lipídios recém-sintetizados. Esse fluxo estabelece a base da via secretora, permitindo que moléculas sejam direcionadas para diferentes destinos celulares ou extracelulares. Em contraposição, o fluxo retrógrado assegura o retorno de componentes essenciais, como enzimas do Golgi e proteínas do retículo, evitando a diluição funcional desses compartimentos. Essa bidirecionalidade confere estabilidade ao sistema, permitindo que ele opere de forma contínua sem perda de eficiência [Tabela].
A integração do sistema também depende da coordenação entre as vias de endocitose e exocitose. A membrana plasmática não é uma estrutura estática, mas um compartimento dinâmico que sofre constante remodelação por meio da incorporação e remoção de vesículas. Material internalizado por endocitose pode ser direcionado para degradação, reciclado para a superfície ou redirecionado para outros compartimentos. Da mesma forma, vesículas exocíticas contribuem para a renovação da membrana e para a liberação de moléculas no meio extracelular. Esse ciclo contínuo estabelece um equilíbrio funcional que regula a composição da superfície celular e sua capacidade de interação com o ambiente.
A comunicação entre compartimentos é mediada por mecanismos moleculares altamente específicos, que garantem o direcionamento correto das vesículas e a fusão seletiva com o compartimento alvo. Proteínas de reconhecimento, pequenas GTPases e complexos de fusão atuam de forma coordenada para assegurar que cada etapa do transporte ocorra com precisão. Essa especificidade é fundamental para evitar a mistura indiscriminada de conteúdos entre organelas, preservando a compartimentalização funcional que caracteriza a célula eucarionte [Quadro].
Outro aspecto central da integração funcional é a existência de gradientes bioquímicos ao longo do sistema de endomembranas. Variações de pH, concentração de íons e composição lipídica criam ambientes distintos que influenciam diretamente a atividade enzimática e o destino das moléculas transportadas. Por exemplo, a acidificação progressiva observada em endossomos e lisossomos não apenas favorece a degradação de biomoléculas, mas também atua como mecanismo de triagem, permitindo a dissociação de complexos moleculares e o direcionamento correto de cargas intracelulares.
A integração do sistema de endomembranas também se estende à sua interação com outros componentes celulares, como o citoesqueleto. Microtúbulos e microfilamentos fornecem as trilhas ao longo das quais vesículas são transportadas, enquanto proteínas motoras garantem o deslocamento direcionado. Essa associação permite que o sistema opere de forma eficiente mesmo em células de grande volume ou altamente polarizadas, nas quais o transporte por difusão seria insuficiente.
Do ponto de vista funcional, o sistema de endomembranas pode ser interpretado como uma rede logística altamente sofisticada, na qual cada compartimento desempenha um papel específico dentro de um fluxo integrado. O retículo endoplasmático atua como centro de produção, o complexo de Golgi como unidade de processamento e triagem, os endossomos como estações de redistribuição e os lisossomos como centros de degradação. A coordenação entre esses elementos garante que a célula mantenha sua organização interna, responda a estímulos externos e adapte-se a mudanças ambientais.
Alterações nesse sistema integrado têm consequências amplas, uma vez que a disfunção em um único componente pode comprometer o fluxo global. Defeitos no transporte vesicular, na fusão de membranas ou na manutenção de gradientes bioquímicos podem resultar em acúmulo de proteínas, falhas na secreção ou degradação inadequada de materiais, evidenciando a dependência mútua entre os compartimentos.
A compreensão da integração funcional do sistema de endomembranas consolida a visão da célula eucarionte como um sistema dinâmico e coordenado, no qual processos de síntese, transporte e degradação estão interligados em uma rede contínua. Essa perspectiva permite avançar além da análise isolada de organelas, enfatizando a importância dos fluxos e das interações que sustentam a funcionalidade celular como um todo.
7.10 — Variações do sistema de endomembranas em diferentes tipos celulares #
A organização do sistema de endomembranas segue princípios estruturais conservados entre células eucariontes, mas sua configuração final não é uniforme. Ao contrário, esse sistema apresenta variações significativas que refletem as demandas funcionais específicas de cada tipo celular. A célula não opera como uma entidade genérica; sua arquitetura interna é moldada pelas exigências metabólicas, fisiológicas e ambientais às quais está submetida. Nesse contexto, o sistema de endomembranas adapta-se estruturalmente para otimizar processos como secreção, absorção, armazenamento e defesa, evidenciando sua plasticidade funcional.
Células especializadas em secreção apresentam um desenvolvimento particularmente acentuado dos componentes associados à via secretora. Nessas células, o retículo endoplasmático rugoso e o complexo de Golgi são altamente expandidos, refletindo a intensa síntese e processamento de proteínas destinadas à exportação. Em células glandulares, por exemplo, observa-se um arranjo polarizado no qual o retículo endoplasmático ocupa regiões próximas ao núcleo, enquanto o Golgi e as vesículas secretoras concentram-se na região voltada para a superfície de secreção. Essa organização espacial favorece o fluxo direcional de moléculas, aumentando a eficiência do processo secretor [Figura].
Em contraste, células com função predominante na detoxificação, como hepatócitos, apresentam uma expansão significativa do retículo endoplasmático liso. Esse compartimento abriga enzimas envolvidas na modificação e neutralização de substâncias potencialmente tóxicas, permitindo que a célula metabolize compostos exógenos e subprodutos metabólicos. A predominância do retículo liso nessas células evidencia como a especialização funcional pode direcionar a reorganização do sistema de endomembranas, priorizando determinadas vias metabólicas em detrimento de outras.
Nas células do sistema imune, o sistema de endomembranas assume características adaptadas à defesa e ao processamento de antígenos. Macrófagos e células dendríticas apresentam sistemas endocíticos altamente desenvolvidos, com abundância de endossomos e lisossomos, refletindo sua capacidade de internalizar, degradar e apresentar moléculas provenientes de patógenos. A eficiência desses processos depende da integração entre endocitose, tráfego vesicular e atividade lisossomal, demonstrando que a organização do sistema de endomembranas está diretamente relacionada à função imunológica dessas células [Tabela].
Em células vegetais, o sistema de endomembranas apresenta características adicionais associadas à presença de vacúolos de grande volume, que desempenham funções de armazenamento, manutenção da pressão osmótica e degradação de materiais. O vacúolo vegetal pode ocupar a maior parte do volume celular, influenciando a distribuição e a organização dos demais compartimentos. Além disso, a interação entre o sistema de endomembranas e organelas específicas, como cloroplastos, introduz novas camadas de complexidade funcional, especialmente no contexto da síntese e do transporte de moléculas relacionadas à fotossíntese.
Células altamente polarizadas, como neurônios, apresentam desafios adicionais para o sistema de endomembranas, uma vez que o transporte intracelular deve ocorrer ao longo de grandes distâncias. Nessas células, o tráfego vesicular depende fortemente da associação com o citoesqueleto, permitindo o deslocamento de vesículas entre o corpo celular e regiões distantes, como axônios e terminações sinápticas. A especialização do sistema de endomembranas em neurônios está diretamente relacionada à necessidade de manter a comunicação rápida e eficiente entre células, destacando a importância da integração entre transporte vesicular e organização estrutural.
A adaptação do sistema de endomembranas também pode ser observada em condições fisiológicas e ambientais variáveis. Alterações na disponibilidade de nutrientes, exposição a agentes tóxicos ou variações na demanda funcional podem induzir mudanças na estrutura e na atividade dos compartimentos membranosos. Essa capacidade de reorganização permite à célula ajustar sua funcionalidade em resposta a estímulos externos, mantendo a homeostase e garantindo sua sobrevivência em diferentes contextos.
Do ponto de vista evolutivo, essas variações refletem a capacidade do sistema de endomembranas de se modular conforme as exigências impostas pela especialização celular. Embora os princípios básicos de organização e funcionamento sejam conservados, a diversidade estrutural observada entre diferentes tipos celulares evidencia que a célula eucarionte opera como um sistema adaptativo, no qual a arquitetura interna é continuamente ajustada para atender a demandas específicas [Quadro].
A compreensão dessas variações permite avançar da visão generalista da célula para uma abordagem mais contextualizada, na qual a organização do sistema de endomembranas é interpretada em função do papel fisiológico da célula no organismo. Essa perspectiva reforça a ideia de que a estrutura celular não é um fim em si mesma, mas um meio pelo qual a função é realizada de forma eficiente e integrada.
Ao integrar essas variações ao modelo geral do sistema de endomembranas, consolida-se a compreensão da célula como uma unidade dinâmica e especializada, capaz de ajustar sua organização interna para sustentar processos biológicos complexos em diferentes níveis de organização.



