6.1 — A célula eucarionte como sistema compartimentalizado #
A passagem da organização procarionte para a organização eucarionte representa uma das transições mais profundas da história celular. No capítulo anterior, a célula procarionte foi apresentada como uma unidade biologicamente eficiente, pequena, metabolicamente versátil e capaz de realizar, em um espaço reduzido, processos essenciais como replicação do DNA, transcrição, tradução, produção de energia e divisão celular. Essa simplicidade, entretanto, não deve ser confundida com limitação biológica. Bactérias e arqueias continuam sendo organismos extraordinariamente bem-sucedidos, capazes de ocupar ambientes extremos, estabelecer relações ecológicas complexas e sustentar ciclos biogeoquímicos fundamentais. O problema surge quando a célula precisa ampliar suas dimensões, aumentar a complexidade de suas reações, regular de modo mais fino sua expressão gênica, distribuir funções no espaço intracelular e coordenar múltiplas atividades simultâneas sem perder controle químico e estrutural. A célula eucarionte surge, nesse contexto, como uma solução arquitetônica: ela não apenas aumenta de tamanho, mas organiza o seu interior em compartimentos especializados, criando uma geografia celular na qual diferentes processos podem ocorrer de maneira simultânea, regulada e integrada. [Figura 6.1]
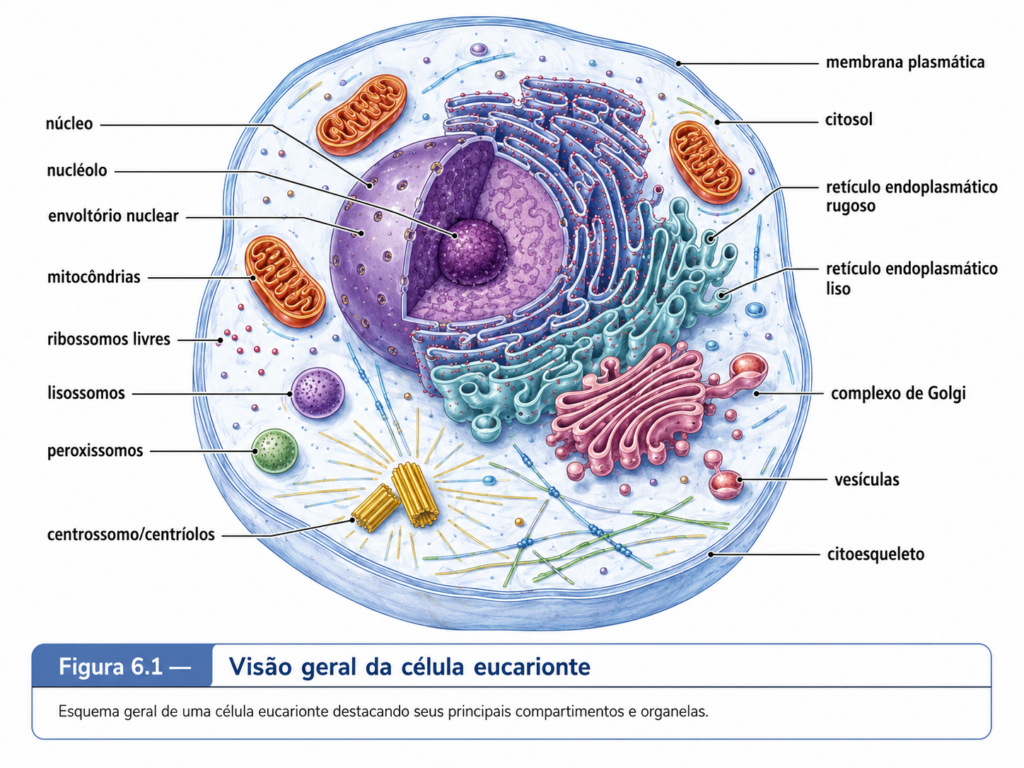
A principal característica estrutural da célula eucarionte é a presença de compartimentos internos delimitados por membranas, entre os quais se destaca o núcleo. Enquanto nas células procariontes o material genético ocupa uma região denominada nucleoide, sem envoltório membranoso próprio, nas células eucariontes o DNA encontra-se separado do citoplasma por um envoltório nuclear. Essa separação permite que processos como transcrição, processamento de RNA e organização da cromatina ocorram em um ambiente próprio, enquanto a tradução proteica se realiza predominantemente no citosol ou associada ao retículo endoplasmático rugoso. A diferença não é apenas morfológica. Na célula procarionte, muitos eventos moleculares ocorrem de forma diretamente acoplada; na célula eucarionte, esses eventos são separados, processados, regulados e reintegrados por mecanismos seletivos de transporte molecular.
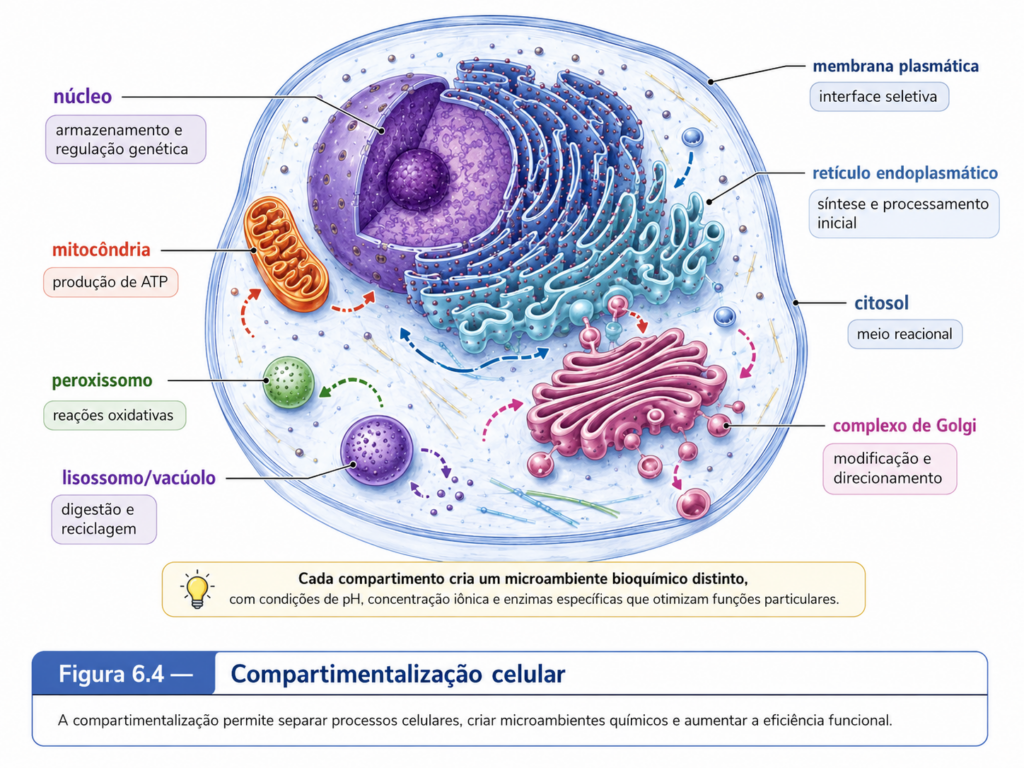
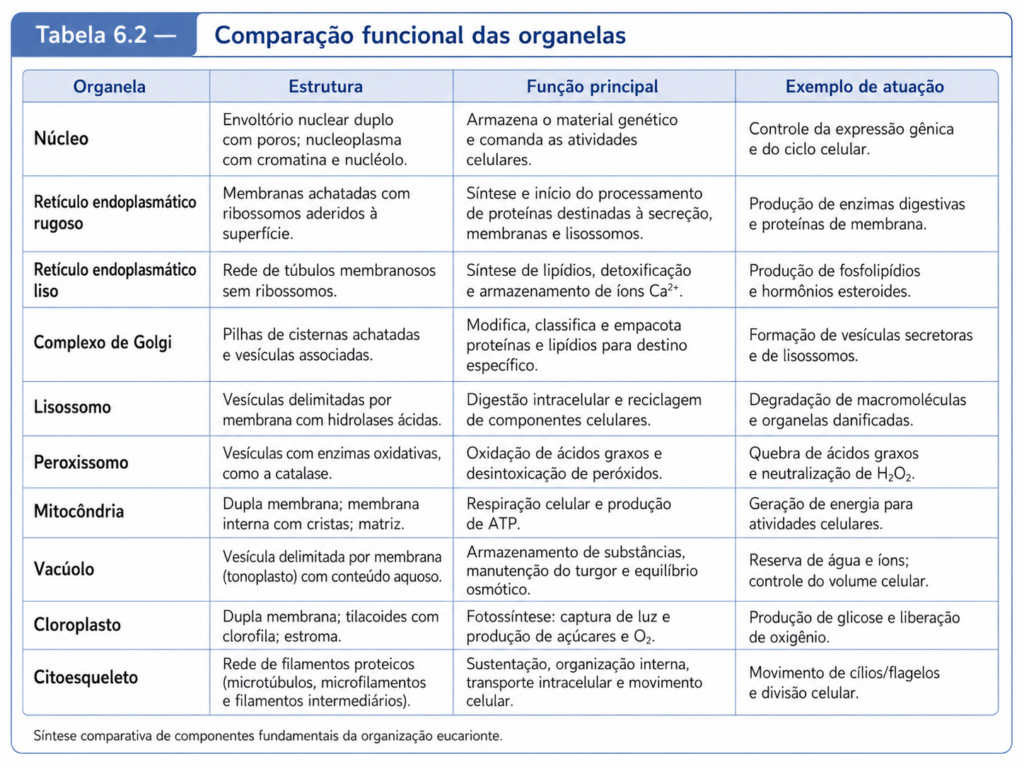
A compartimentalização não deve ser entendida apenas como a presença de organelas dentro da célula. Ela representa uma estratégia de organização bioquímica. Cada compartimento celular possui composição molecular própria, conjunto específico de enzimas, pH característico, gradientes iônicos, proteínas residentes, sistemas de transporte e funções predominantes. O retículo endoplasmático participa da síntese e do processamento inicial de proteínas e lipídios; o complexo de Golgi modifica, seleciona e direciona moléculas; os lisossomos e vacúolos atuam na digestão intracelular e na reciclagem de componentes; as mitocôndrias participam da conversão energética; os cloroplastos, nas células vegetais e algas, capturam energia luminosa e a convertem em energia química; os peroxissomos realizam reações oxidativas importantes; e o núcleo preserva, organiza e regula a informação genética. [Tabela 6.1]
Essa organização permite que reações potencialmente incompatíveis ocorram dentro da mesma célula sem interferência direta. Enzimas digestivas, por exemplo, poderiam degradar componentes celulares se estivessem dispersas livremente no citosol; ao serem concentradas em lisossomos ou vacúolos, tornam-se úteis sem comprometer a integridade celular. Da mesma forma, reações oxidativas que geram espécies reativas de oxigênio podem ser confinadas em peroxissomos ou mitocôndrias, onde sistemas antioxidantes específicos limitam seus efeitos colaterais. A célula eucarionte, portanto, não resolve o problema da complexidade apenas produzindo mais moléculas. Ela resolve esse problema distribuindo funções no espaço.
A membrana é o elemento central dessa lógica. Como visto no capítulo sobre membranas celulares, a bicamada lipídica não atua somente como barreira física. Ela cria superfícies seletivas, delimita ambientes químicos, organiza proteínas, sustenta gradientes, orienta fluxos moleculares e permite comunicação entre compartimentos. Na célula eucarionte, essa propriedade é expandida ao máximo. A membrana plasmática separa a célula do meio externo, enquanto as membranas internas subdividem o citoplasma em regiões funcionais. A consequência é que o interior celular deixa de ser um espaço homogêneo e passa a funcionar como um conjunto integrado de microambientes bioquímicos.
A consolidação dessa visão dependeu de uma mudança técnica decisiva na história da biologia celular. A microscopia óptica havia permitido reconhecer células, núcleos e algumas estruturas maiores, mas a organização fina do citoplasma permaneceu inacessível até o desenvolvimento e a aplicação ampla da microscopia eletrônica. Com ela, tornou-se possível observar envoltório nuclear, retículo endoplasmático, complexo de Golgi, mitocôndrias, cloroplastos, lisossomos, peroxissomos e vesículas com um nível de resolução incompatível com os instrumentos anteriores. Ainda assim, ver estruturas não bastava. Era necessário demonstrar que essas estruturas possuíam funções próprias. O fracionamento celular, especialmente por centrifugação diferencial, permitiu romper células cuidadosamente e separar componentes subcelulares em frações distintas, associando determinadas estruturas a atividades bioquímicas específicas. A combinação entre imagem e bioquímica transformou a célula eucarionte de uma entidade apenas observável em um sistema funcionalmente mapeável.
Essa arquitetura compartimentalizada favoreceu uma elevação expressiva da eficiência funcional. A célula eucarionte pode concentrar substratos, enzimas e produtos em regiões específicas, reduzir interferências entre vias metabólicas, controlar fluxos moleculares e criar ambientes internos especializados. Em uma célula secretora, proteínas destinadas à exportação são sintetizadas no retículo endoplasmático rugoso, modificadas no complexo de Golgi, empacotadas em vesículas e direcionadas à membrana plasmática. Em uma célula vegetal fotossintética, a produção de açúcares nos cloroplastos precisa ser coordenada com a respiração mitocondrial, o metabolismo citosólico, o armazenamento vacuolar e o transporte de solutos entre tecidos. Em ambos os casos, a função celular emerge da integração entre compartimentos, não da ação isolada de uma organela.
Nas plantas submetidas a déficit hídrico, salinidade ou calor excessivo, essa integração se torna particularmente evidente. O vacúolo participa do ajuste osmótico e do armazenamento de íons; os cloroplastos ajustam a atividade fotossintética conforme a disponibilidade de luz, água e CO₂; as mitocôndrias reorganizam o metabolismo respiratório; os peroxissomos participam de reações oxidativas e da contenção de espécies reativas de oxigênio; as membranas modulam transporte e sinalização; e o núcleo altera padrões de expressão gênica em resposta ao estresse. A tolerância de uma cultivar à seca ou à salinidade não depende de uma única estrutura celular, mas da coordenação entre múltiplos compartimentos. Esse entendimento sustenta estratégias modernas de melhoramento vegetal, bioestimulação, manejo nutricional e biotecnologia voltadas à estabilidade produtiva em ambientes limitantes. [Quadro 6.1]
A compartimentalização também ampliou a capacidade regulatória da célula. Ao separar processos em compartimentos distintos, a célula pode controlar não apenas se uma reação ocorrerá, mas onde, quando e em que intensidade ela ocorrerá. Uma proteína recém-sintetizada pode permanecer no citosol, entrar no núcleo, ser enviada ao retículo endoplasmático, incorporada a uma membrana, transportada para uma mitocôndria ou direcionada à degradação. Esse destino depende de sinais moleculares presentes na própria proteína, de receptores específicos, de sistemas de transporte, de vesículas e de mecanismos de reconhecimento intracelular. A célula eucarionte, portanto, não é apenas compartimentalizada; ela é endereçada. Suas moléculas carregam informações de localização, e seus compartimentos possuem mecanismos seletivos de entrada, retenção e exportação.
Essa organização tem consequências diretas para a compreensão da vida multicelular. Tecidos, órgãos e organismos complexos só são possíveis porque suas células conseguem especializar funções mantendo uma organização interna altamente controlada. Uma célula muscular depende da coordenação entre retículo sarcoplasmático, mitocôndrias, citoesqueleto e membrana plasmática para contrair. Uma célula glandular depende do sistema de endomembranas para secretar. Uma célula meristemática vegetal depende da integração entre núcleo, citosol, vacúolo, parede celular e citoesqueleto para crescer e dividir-se. Uma célula do mesófilo foliar depende da relação funcional entre cloroplastos, mitocôndrias, peroxissomos e vacúolo para sustentar fotossíntese, respiração, fotorrespiração, controle redox e balanço hídrico. [Figura 6.2]
Um erro comum no estudo inicial de biologia celular é tratar a célula eucarionte como uma coleção de peças isoladas. Essa abordagem facilita a memorização, mas prejudica a compreensão. O núcleo não funciona isoladamente do citoplasma; as mitocôndrias não operam sem intercâmbio de metabólitos e proteínas codificadas no núcleo; o retículo endoplasmático não atua sem relação com o complexo de Golgi, vesículas, membrana plasmática e sistemas de degradação; o citoesqueleto não é apenas sustentação, mas um sistema de organização espacial, transporte e resposta mecânica. A célula viva não se comporta como um conjunto de organelas justapostas, mas como uma rede dinâmica de compartimentos interdependentes.
A origem dessa arquitetura ainda é tema de investigação e debate científico, mas dois processos são fundamentais para compreendê-la. O primeiro envolve a formação progressiva de membranas internas, possivelmente associada a invaginações da membrana plasmática ancestral e à especialização de domínios membranosos. Esse processo ajuda a explicar a origem do sistema de endomembranas, incluindo envoltório nuclear, retículo endoplasmático, complexo de Golgi, endossomos, lisossomos e vesículas. O segundo envolve eventos endossimbióticos, nos quais ancestrais de mitocôndrias e cloroplastos teriam derivado de bactérias incorporadas por células ancestrais e mantidas ao longo da evolução por sua contribuição energética e metabólica. Essa interpretação é sustentada por características como dupla membrana, DNA próprio, ribossomos próprios e divisão por processos relacionados à fissão. [Figura 6.3]
A vantagem evolutiva da compartimentalização não foi apenas permitir que células maiores existissem. O ganho mais profundo foi permitir que a célula desenvolvesse níveis adicionais de controle. Células maiores enfrentam desafios físicos importantes, como maiores distâncias internas para difusão, aumento da demanda energética e necessidade de transporte dirigido. A organização eucarionte responde a esses desafios com membranas internas, organelas, citoesqueleto, motores moleculares, poros nucleares, vesículas, sinais de endereçamento e sistemas de comunicação intracelular. Assim, o aumento de tamanho torna-se biologicamente viável porque a célula deixa de depender apenas da difusão aleatória e passa a utilizar mecanismos organizados de distribuição, confinamento e transporte.
Nas células vegetais, essa lógica atinge uma expressão particularmente relevante para as ciências agrárias. Além das estruturas comuns às células eucariontes, essas células possuem parede celular, grandes vacúolos, plastídios e plasmodesmos. O vacúolo central participa do equilíbrio osmótico, do armazenamento de íons e metabólitos, da degradação de componentes celulares e da manutenção do turgor. Os cloroplastos permitem a fixação de carbono, mas sua atividade precisa ser ajustada à disponibilidade de luz, água, CO₂ e nutrientes. A parede celular impõe rigidez, mas também participa do crescimento, da defesa e da comunicação entre células. Em condições de estresse salino, hídrico ou térmico, a sobrevivência da planta depende da coordenação entre compartimentos celulares, e não da ação isolada de uma única organela.
Essa visão é essencial para interpretar fenômenos biotecnológicos e fisiológicos. A produtividade agrícola, a tolerância à seca, a eficiência fotossintética, a absorção de nutrientes, a resistência a patógenos e a qualidade pós-colheita dependem de processos que ocorrem em múltiplos compartimentos celulares. Uma mutação que altera uma proteína de membrana pode modificar transporte iônico; uma deficiência em enzimas peroxissomais pode comprometer o metabolismo oxidativo; uma alteração na dinâmica do citoesqueleto pode afetar expansão celular; uma falha no tráfego vesicular pode prejudicar secreção de componentes da parede celular; uma disfunção mitocondrial pode alterar energia, sinalização e morte celular. A célula eucarionte deve, portanto, ser estudada como um sistema regulado em rede.
A compreensão da compartimentalização também modifica a forma como se interpreta a expressão gênica. O DNA nuclear contém grande parte das instruções para a produção de proteínas celulares, mas a execução dessas instruções depende do citoplasma e das organelas. Muitas proteínas que atuam em mitocôndrias, cloroplastos, peroxissomos e no sistema de endomembranas são codificadas no núcleo, sintetizadas por ribossomos citosólicos e depois direcionadas ao compartimento correto. Isso significa que a informação genética não basta por si só: ela precisa ser transcrita, processada, traduzida, endereçada, importada, modificada e integrada a estruturas funcionais. A célula eucarionte é, portanto, uma arquitetura de informação, matéria e energia.
Ao final, a célula eucarionte não pode ser definida apenas pela presença de núcleo. O núcleo é uma marca estrutural decisiva, mas a identidade eucariótica depende de um conjunto integrado de características: compartimentalização membranosa, citosol organizado, organelas especializadas, citoesqueleto dinâmico, tráfego intracelular, separação entre transcrição e tradução, controle de endereçamento molecular e comunicação entre compartimentos. Essa organização cria as condições para a especialização celular, para a multicelularidade complexa e para a diversidade funcional observada em animais, plantas, fungos, algas e protozoários. Nos próximos níveis de análise, cada um desses componentes poderá ser examinado com maior detalhe, mas a chave interpretativa permanece a mesma: a célula eucarionte é uma unidade integrada cuja complexidade resulta da cooperação espacial e funcional entre seus compartimentos.
6.2 — Citoplasma, citosol e organização interna da célula #
A compreensão da célula eucarionte exige uma distinção inicial que, embora pareça simples, evita muitos erros conceituais: citoplasma e citosol não são sinônimos. O citoplasma corresponde a todo o conteúdo celular situado entre a membrana plasmática e o núcleo, incluindo organelas, vesículas, citoesqueleto, inclusões, ribossomos e a fase aquosa que envolve esses componentes. O citosol, por sua vez, é a porção fluida e não delimitada por membranas do citoplasma, isto é, o meio intracelular no qual se encontram dispersos íons, metabólitos, proteínas solúveis, RNAs, ribossomos livres, enzimas, complexos macromoleculares e estruturas associadas ao citoesqueleto. Essa diferença é fundamental porque o citoplasma inclui compartimentos membranosos especializados, enquanto o citosol representa o ambiente contínuo que conecta grande parte da atividade molecular da célula. [Figura 6.4]
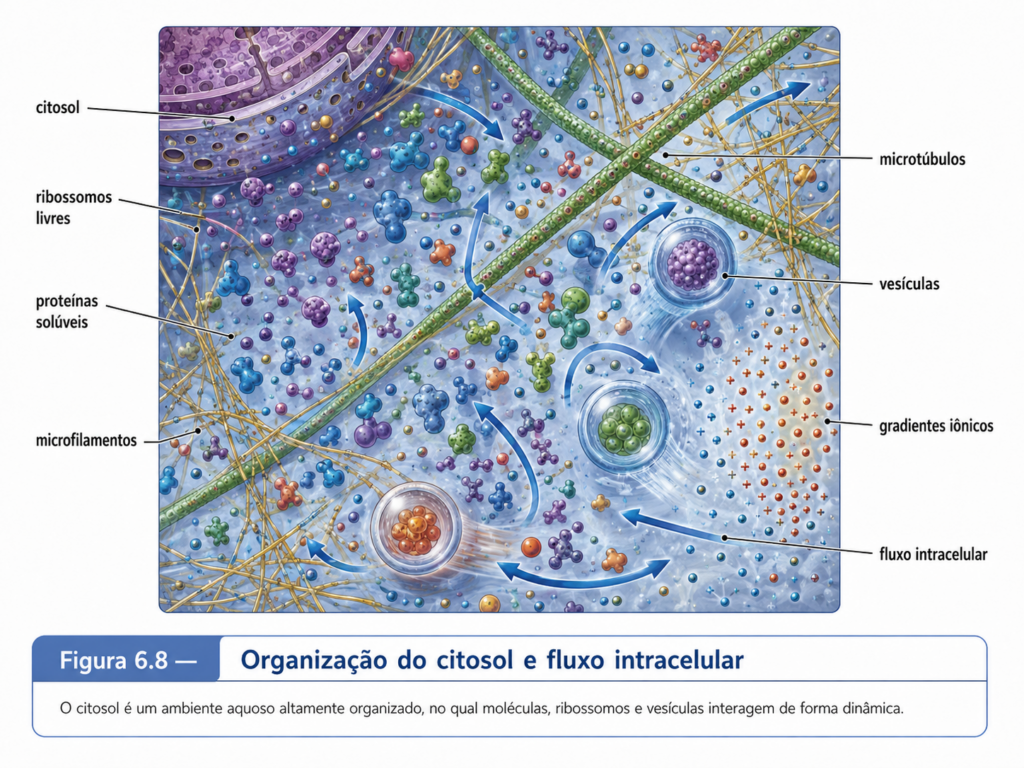
Durante muito tempo, o citosol foi imaginado de maneira simplificada, como se fosse apenas uma solução aquosa na qual organelas estariam suspensas. Essa imagem é didaticamente confortável, mas biologicamente limitada. O citosol não é um líquido diluído semelhante à água pura. Trata-se de um meio altamente concentrado, viscoso, dinâmico e densamente ocupado por moléculas de diferentes tamanhos. Proteínas, RNAs, ribossomos, metabólitos e íons não estão distribuídos em um vazio molecular, mas em um ambiente no qual o espaço disponível é disputado continuamente. Essa elevada concentração molecular altera a difusão, favorece encontros entre moléculas compatíveis, influencia o dobramento de proteínas, modifica equilíbrios químicos e cria condições para que reações metabólicas ocorram em velocidades adequadas à vida celular. [Figura 6.5]
A água continua sendo o componente predominante do citosol, mas não se comporta ali como um solvente indiferente. Parte das moléculas de água interage com proteínas, ácidos nucleicos, íons e superfícies carregadas, formando camadas de hidratação essenciais para a estabilidade estrutural das macromoléculas. Outra parte permanece mais livre, permitindo difusão e reações químicas. Essa organização faz com que o citosol funcione como uma matriz aquosa viva, na qual as propriedades da água estudadas no capítulo sobre química celular reaparecem em um nível mais complexo. Polaridade, ligações de hidrogênio, interações iônicas, efeito hidrofóbico e equilíbrio ácido-base deixam de ser abstrações químicas e passam a sustentar a atividade metabólica concreta da célula.
No citosol encontram-se muitos dos metabólitos intermediários que conectam vias biossintéticas, energéticas e regulatórias. A glicólise, por exemplo, ocorre no citosol e representa uma das vias centrais de degradação da glicose, fornecendo ATP, NADH e intermediários que alimentam outras rotas metabólicas. A via das pentoses fosfato também ocorre nesse ambiente, produzindo NADPH e açúcares fosfatados importantes para biossíntese e equilíbrio redox. Aminoácidos, nucleotídeos, açúcares, ácidos orgânicos, coenzimas e pequenas moléculas regulatórias circulam pelo citosol e fazem dele um centro de integração metabólica. Ainda que mitocôndrias, cloroplastos e peroxissomos concentrem etapas especializadas do metabolismo, o citosol permanece como uma plataforma de conexão entre essas organelas e o restante da célula.
Essa integração é particularmente evidente quando se observa o metabolismo energético. Em células animais, a glicose pode ser degradada inicialmente no citosol, gerando piruvato, que será posteriormente importado pelas mitocôndrias para alimentar o ciclo do ácido cítrico e a fosforilação oxidativa. Em células vegetais, açúcares produzidos ou mobilizados a partir de processos relacionados aos cloroplastos podem ser metabolizados no citosol, armazenados em vacúolos, convertidos em sacarose ou direcionados a diferentes tecidos. O citosol, portanto, não é um compartimento residual entre organelas; ele é o espaço onde rotas metabólicas se encontram, onde decisões bioquímicas são ajustadas e onde a célula distribui carbono, energia e poder redutor conforme suas necessidades fisiológicas. [Tabela 6.2]
Além de abrigar vias metabólicas, o citosol é o principal local de síntese de proteínas por ribossomos livres. Todos os ribossomos eucarióticos iniciam a tradução no citosol, mas o destino da proteína nascente depende de sinais moleculares presentes em sua sequência. Proteínas que permanecerão no citosol, entrarão no núcleo, serão importadas para mitocôndrias, cloroplastos ou peroxissomos, ou ainda seguirão para o sistema de endomembranas, começam sua trajetória nesse mesmo ambiente. Quando a proteína possui sinal adequado para o retículo endoplasmático, a tradução é redirecionada para ribossomos associados à membrana do retículo. Quando não possui esse sinal, a tradução pode prosseguir em ribossomos livres, produzindo proteínas destinadas ao próprio citosol ou a outros compartimentos por importação pós-traducional. Dessa forma, o citosol é simultaneamente local de produção, triagem inicial e distribuição potencial de grande parte das proteínas celulares.
Essa condição torna o citosol um ponto estratégico para o controle da informação genética. O núcleo armazena o DNA e realiza a transcrição, mas muitas consequências funcionais da expressão gênica se concretizam no citosol, onde mRNAs são traduzidos, degradados, silenciados ou direcionados a regiões específicas da célula. A estabilidade de um mRNA, sua taxa de tradução, sua associação com ribossomos e sua localização citoplasmática influenciam diretamente a quantidade e o local de produção de uma proteína. Em células polarizadas, como neurônios, células epiteliais ou células vegetais em crescimento, a localização de RNAs e proteínas no citoplasma permite respostas espacialmente controladas. A célula não apenas produz moléculas; ela as posiciona.
O citosol também abriga sistemas de controle de qualidade molecular. Proteínas recém-sintetizadas precisam dobrar-se corretamente, muitas vezes com auxílio de chaperonas moleculares. Quando esse dobramento falha, proteínas mal conformadas podem ser reconhecidas, marcadas e degradadas por sistemas como o ubiquitina-proteassoma. Esse controle evita o acúmulo de agregados tóxicos e mantém a funcionalidade do proteoma celular. A vida no citosol, portanto, envolve uma tensão permanente entre síntese, dobramento, modificação, uso e degradação de proteínas. A célula precisa produzir moléculas suficientes para sustentar sua atividade, mas também precisa remover aquelas que envelhecem, perdem função ou se tornam perigosas.
A concentração de íons no citosol é rigidamente controlada. Potássio, magnésio, sódio, cloreto, fosfato e cálcio participam de processos estruturais, osmóticos, enzimáticos e sinalizadores. Entre eles, o cálcio ocupa posição especial. A concentração de Ca²⁺ livre no citosol é mantida em níveis muito baixos quando comparada ao meio extracelular e ao interior de certos compartimentos, como o retículo endoplasmático. Pequenos aumentos na concentração citosólica de Ca²⁺ podem atuar como sinais capazes de modificar atividade enzimática, secreção, contração, metabolismo, expressão gênica e respostas ao estresse. Essa propriedade só é possível porque a célula mantém gradientes iônicos por meio de bombas, canais e transportadores localizados na membrana plasmática e em membranas internas. O citosol, nesse caso, funciona como espaço de leitura de sinais, no qual alterações transitórias de íons podem ser convertidas em respostas bioquímicas amplificadas. [Animação 6.1]
A regulação do pH citosólico é igualmente importante. Muitas enzimas possuem atividade ótima em faixas específicas de pH, e alterações acentuadas podem comprometer reações metabólicas, estrutura proteica e transporte de moléculas. A célula mantém o pH citosólico por meio de sistemas tampão, transportadores de prótons, trocadores iônicos e interação com organelas capazes de armazenar ou liberar íons. Essa estabilidade não significa ausência de variação. Em determinadas condições fisiológicas, mudanças locais ou transitórias de pH podem regular processos específicos. Ainda assim, a manutenção de um ambiente citosólico compatível com a atividade enzimática é uma condição indispensável para a homeostase celular.
A ideia de que o citosol é homogêneo também vem sendo superada pela compreensão de sua organização espacial. Mesmo sem membranas delimitando todas as regiões internas, o citosol contém zonas funcionais, gradientes locais, complexos proteicos, agrupamentos ribonucleoproteicos e estruturas transitórias. Algumas enzimas de uma mesma via metabólica podem associar-se fisicamente, favorecendo a transferência eficiente de intermediários entre etapas sucessivas. Determinados RNAs e proteínas podem concentrar-se em regiões específicas sem que exista uma membrana ao redor. Em muitos casos, interações fracas e reversíveis entre moléculas permitem a formação de condensados biomoleculares, estruturas dinâmicas que concentram componentes relacionados a determinadas funções. Essa organização mostra que a compartimentalização celular não depende apenas de membranas; ela também pode emergir de interações moleculares seletivas dentro do próprio citosol.
O citoesqueleto contribui decisivamente para essa organização interna. Filamentos de actina, microtúbulos e filamentos intermediários atravessam o citoplasma e conferem suporte, forma, polaridade e capacidade de movimento à célula. Embora o citoesqueleto seja tratado com maior profundidade em capítulo posterior, sua presença precisa ser reconhecida desde já porque o citosol não pode ser compreendido como uma massa desorganizada. Microtúbulos podem orientar o transporte de vesículas e organelas; filamentos de actina participam de movimentos corticais, contração, expansão celular e organização da periferia; filamentos intermediários contribuem para resistência mecânica em muitos tipos celulares. A estrutura citosólica é, portanto, simultaneamente química e física: moléculas reagem, mas também se distribuem, se ancoram, se deslocam e respondem a forças.
A movimentação intracelular depende dessa relação entre citosol e citoesqueleto. Vesículas, organelas, complexos proteicos e RNAs podem deslocar-se por difusão, mas muitos movimentos celulares exigem transporte dirigido. Proteínas motoras, como cinesinas, dineínas e miosinas, convertem energia química em trabalho mecânico e transportam cargas ao longo de microtúbulos ou filamentos de actina. Esse transporte evita que a célula dependa exclusivamente da difusão, especialmente em células grandes ou polarizadas. Em uma célula eucarionte, a distância entre o local de síntese, modificação, uso e degradação de uma molécula pode ser significativa; por isso, a organização do citosol como meio estruturado e atravessado por rotas de transporte é essencial para a eficiência celular.
Nas células vegetais, essa dinâmica assume uma expressão particularmente visível no movimento citoplasmático, também chamado ciclose. Em muitas células vegetais, correntes citoplasmáticas deslocam organelas e componentes internos ao longo de trajetórias associadas ao citoesqueleto, favorecendo distribuição de metabólitos, posicionamento de cloroplastos, comunicação entre regiões celulares e ajuste funcional em células de grande volume. Quando o vacúolo central ocupa a maior parte do espaço interno, o citosol pode ficar comprimido em uma camada periférica fina entre a membrana plasmática e o tonoplasto, além de formar cordões citoplasmáticos que atravessam a célula. Mesmo reduzido em volume relativo, esse citosol mantém intensa atividade metabólica e organizacional, reforçando que sua importância não depende apenas da quantidade, mas da função estratégica que exerce. [Figura 6.6]
A relação entre citosol e vacúolo é central para a fisiologia vegetal. O vacúolo armazena íons, metabólitos, pigmentos, compostos de defesa e água, enquanto o citosol mantém as reações metabólicas sensíveis a variações químicas. Para que essa separação funcione, o tonoplasto controla fluxos entre os dois ambientes, permitindo que a célula ajuste seu potencial osmótico sem desorganizar completamente o metabolismo citosólico. Em situações de seca ou salinidade, por exemplo, plantas podem acumular solutos compatíveis no citosol e compartimentalizar íons no vacúolo, reduzindo efeitos tóxicos e preservando atividade enzimática. A tolerância celular ao estresse depende, nesse caso, da capacidade de manter o citosol funcional enquanto outros compartimentos absorvem parte da instabilidade imposta pelo ambiente. [Quadro 6.2]
A organização do citosol também é decisiva para a sinalização celular. Muitas vias de sinalização começam na membrana plasmática, quando receptores percebem hormônios, nutrientes, patógenos, luz, pressão mecânica ou alterações ambientais. O sinal inicial frequentemente precisa ser transmitido ao citosol por segundos mensageiros, proteínas quinases, fosfatases, pequenas GTPases, alterações iônicas ou mudanças no estado redox. A partir daí, a informação pode modificar enzimas citosólicas, reorganizar o citoesqueleto, alterar o tráfego vesicular ou alcançar o núcleo, onde influencia a expressão gênica. Assim, o citosol atua como meio de propagação, integração e filtragem de sinais. Ele não apenas recebe informações; ele decide, por meio de redes moleculares, quais respostas serão amplificadas, atenuadas ou combinadas.
Essa função integradora é evidente na resposta de plantas a estresses ambientais. Sob déficit hídrico, por exemplo, a percepção da redução de água disponível desencadeia mudanças em hormônios vegetais, fluxos iônicos, espécies reativas de oxigênio, concentração de Ca²⁺ citosólico, metabolismo de solutos compatíveis e expressão gênica. O citosol participa desse processo como espaço de convergência entre sinais vindos da membrana plasmática, do vacúolo, dos cloroplastos, das mitocôndrias e dos peroxissomos. Quando a intensidade luminosa excede a capacidade de uso da energia capturada pelos cloroplastos, alterações redox e espécies reativas podem influenciar respostas citosólicas e nucleares. Quando a salinidade aumenta, transportadores ajustam fluxos de Na⁺, K⁺ e H⁺, e o citosol precisa preservar enzimas e estruturas sensíveis à perturbação iônica. O desempenho fisiológico da planta emerge dessa rede de ajustes, e não de uma resposta isolada.
A própria noção de metabolismo celular torna-se mais precisa quando se reconhece o citosol como ambiente organizado. Vias metabólicas não são sequências abstratas de reações desenhadas em esquemas lineares; elas ocorrem em espaços físicos, dependem da concentração local de substratos, da disponibilidade de cofatores, da proximidade entre enzimas, do estado energético da célula e da comunicação com organelas. A glicólise, a síntese de ácidos graxos em determinados contextos, parte do metabolismo de aminoácidos, a ativação de açúcares e numerosas reações de sinalização ocorrem em um ambiente no qual cada molécula compete por espaço, interage com superfícies e responde a mudanças rápidas. A célula não é um tubo de ensaio ampliado; é um sistema espacialmente organizado em escala microscópica e molecular.
Os avanços metodológicos foram decisivos para essa mudança de interpretação. A microscopia revelou que o citoplasma não era vazio nem uniforme, enquanto o fracionamento celular permitiu separar componentes e associar determinadas atividades bioquímicas a frações específicas. A ultracentrifugação diferencial, ao separar núcleos, mitocôndrias, microssomos, ribossomos e frações solúveis, ajudou a distinguir o que pertencia a organelas membranosas e o que permanecia na fração citosólica. Posteriormente, técnicas de marcação fluorescente, microscopia confocal, recuperação de fluorescência após fotobranqueamento, rastreamento de moléculas individuais e proteômica ampliaram a percepção de que o citosol é dinâmico, heterogêneo e regulado. A célula passou a ser interpretada não apenas como uma imagem fixa, mas como um sistema de fluxos, interações e reorganizações constantes.
Essa perspectiva também corrige uma limitação frequente no ensino introdutório: a tendência de valorizar organelas visíveis em esquemas e subestimar o espaço entre elas. Como núcleo, mitocôndrias, cloroplastos, retículo endoplasmático e Golgi possuem formas reconhecíveis, tornam-se mais fáceis de representar e memorizar. O citosol, por não ter contorno próprio, parece menos importante. No entanto, grande parte da vida molecular da célula ocorre justamente nesse ambiente. É nele que proteínas são sintetizadas, muitas enzimas atuam, sinais se propagam, metabólitos circulam, complexos se formam, moléculas são degradadas e organelas se comunicam. Ignorar o citosol equivale a descrever uma cidade apenas por seus prédios, sem considerar as ruas, redes, fluxos, pessoas e sistemas que fazem a cidade funcionar.
A diferença entre citosol e organelas também não deve sugerir separação absoluta. Mitocôndrias importam proteínas sintetizadas no citosol e exportam metabólitos que alimentam vias citosólicas. Cloroplastos dependem de proteínas codificadas no núcleo e sintetizadas por ribossomos citosólicos. Peroxissomos recebem proteínas produzidas no citosol e participam de rotas que envolvem lipídios, espécies reativas de oxigênio e metabolismo vegetal. O retículo endoplasmático interage com o citosol tanto na síntese proteica quanto na regulação de cálcio e lipídios. O núcleo exporta RNAs para o citosol e importa proteínas regulatórias a partir dele. A vida eucariótica depende dessa troca permanente entre compartimentos delimitados por membranas e o ambiente citosólico que os conecta.
Em células animais, alterações citosólicas podem estar associadas a processos patológicos relevantes. Distúrbios no dobramento proteico, falhas na degradação pelo proteassoma, alterações em vias de sinalização, desequilíbrios de Ca²⁺, mudanças no pH intracelular e disfunções do citoesqueleto podem afetar sobrevivência, proliferação, secreção, migração e morte celular. Em células vegetais, problemas na manutenção do equilíbrio osmótico, na organização do citoesqueleto, no controle redox e na comunicação entre citosol e organelas podem comprometer crescimento, produtividade e tolerância a estresses. A análise do citosol, portanto, não é apenas um detalhe da biologia celular básica; ela é necessária para compreender fisiologia, patologia, biotecnologia e adaptação.
O citoplasma, visto em sua totalidade, representa o espaço operacional da célula eucarionte. Nele, organelas delimitadas por membranas executam funções especializadas, enquanto o citosol conecta essas funções por meio de reações químicas, difusão, transporte dirigido, sinalização, síntese proteica e controle molecular. Essa organização permite que a célula seja compartimentalizada sem se tornar fragmentada. O citosol mantém continuidade funcional entre estruturas distintas, funcionando como matriz de integração metabólica, informacional e mecânica. A célula eucarionte, portanto, não deve ser imaginada como um conjunto de organelas flutuando em um líquido indiferente, mas como um sistema interno altamente estruturado, no qual o citosol exerce papel central na coordenação da vida molecular.
6.3 — Organelas membranosas e especialização funcional #
A célula eucarionte não se tornou mais complexa apenas porque acumulou mais moléculas em seu interior. O salto organizacional ocorreu quando parte dessas moléculas passou a ser reunida em compartimentos delimitados por membranas, cada um com composição, propriedades físico-químicas e funções predominantes. Essa compartimentalização permitiu que a célula separasse processos incompatíveis, concentrasse enzimas e substratos, controlasse fluxos de matéria e energia, protegesse componentes sensíveis e estabelecesse circuitos internos de comunicação. As organelas membranosas representam, portanto, uma das expressões mais sofisticadas da organização eucariótica: são espaços internos especializados, mas interdependentes, nos quais a célula distribui trabalho molecular sem perder unidade funcional. [Figura 6.7]
A noção de organela membranosa deve ser entendida com cuidado. Uma organela não é apenas uma estrutura desenhada em esquemas didáticos, com forma fixa e função isolada. Ela é um compartimento dinâmico, delimitado por uma ou mais membranas, contendo proteínas residentes, sistemas de transporte, enzimas específicas, gradientes químicos e relações funcionais com outros compartimentos. Algumas organelas, como núcleo, retículo endoplasmático, complexo de Golgi, endossomos, lisossomos e vacúolos, integram o sistema de endomembranas ou se relacionam diretamente com ele. Outras, como mitocôndrias e cloroplastos, possuem origem evolutiva distinta, apresentam dupla membrana, material genético próprio e importam grande parte de suas proteínas a partir do citosol. Os peroxissomos, por sua vez, combinam simplicidade morfológica aparente com grande importância metabólica, especialmente em reações oxidativas e na integração com lipídios, espécies reativas de oxigênio e metabolismo vegetal. [Tabela 6.3]
A membrana é o princípio organizador de todas essas estruturas. Ao delimitar um compartimento, a membrana cria diferença. De um lado, estabelece uma composição interna; de outro, mantém relação seletiva com o citosol. Essa separação permite que o interior de uma organela tenha pH, concentração iônica, conjunto enzimático e estado redox distintos do ambiente citosólico. Em lisossomos e vacúolos, por exemplo, a acidificação interna favorece a atividade de enzimas hidrolíticas. Em mitocôndrias e cloroplastos, membranas internas sustentam cadeias de transporte de elétrons e gradientes eletroquímicos envolvidos na síntese de ATP. No retículo endoplasmático, a membrana delimita um lúmen especializado em dobramento, modificação e controle de qualidade de proteínas destinadas à secreção, à membrana plasmática ou a compartimentos internos. No complexo de Golgi, pilhas de cisternas membranosas criam uma sequência de ambientes nos quais proteínas e lipídios são modificados, selecionados e direcionados.
A especialização funcional das organelas depende, em grande medida, da composição de suas membranas. A bicamada lipídica fornece a matriz estrutural, mas são as proteínas de membrana que determinam grande parte das propriedades específicas de cada compartimento. Bombas, canais, transportadores, receptores, enzimas, proteínas de ancoragem, proteínas de fusão vesicular e complexos de translocação tornam as membranas superfícies funcionais. Uma membrana mitocondrial interna, rica em proteínas relacionadas ao transporte de elétrons e à síntese de ATP, tem propriedades muito diferentes da membrana do retículo endoplasmático, envolvida na translocação de proteínas, síntese lipídica e armazenamento de cálcio. O mesmo princípio vale para o tonoplasto vegetal, cuja composição permite ao vacúolo atuar na regulação osmótica, armazenamento e degradação, e para a membrana dos peroxissomos, que controla a entrada de proteínas e metabólitos envolvidos em reações oxidativas.
Essa diversidade membranosa permite que a célula transforme espaço em regulação. Uma reação química não depende apenas da presença de enzima e substrato; depende também de localização, concentração, acesso, retenção e remoção de produtos. Ao concentrar determinadas enzimas em organelas específicas, a célula aumenta a eficiência de vias metabólicas e reduz interferências. Enzimas hidrolíticas são mantidas em compartimentos digestivos; enzimas da respiração celular são associadas a membranas e matrizes mitocondriais; enzimas da fotossíntese são organizadas em tilacoides e estroma dos cloroplastos; enzimas oxidativas são concentradas em peroxissomos; enzimas de modificação de proteínas atuam em regiões específicas do retículo endoplasmático e do Golgi. A organela, nesse sentido, não é um recipiente passivo. Ela é uma unidade de condições químicas controladas.
O retículo endoplasmático ilustra bem essa relação entre estrutura e função. Sua rede de membranas se estende por grande parte do citoplasma e se comunica estruturalmente com o envoltório nuclear. Em regiões associadas a ribossomos, forma o retículo endoplasmático rugoso, relacionado à síntese de proteínas secretadas, proteínas de membrana e proteínas destinadas a compartimentos do sistema de endomembranas. Em regiões sem ribossomos aderidos, forma o retículo endoplasmático liso, importante para síntese de lipídios, metabolismo de substâncias hidrofóbicas e armazenamento de cálcio em determinados tipos celulares. Essa divisão entre rugoso e liso não representa dois órgãos separados, mas domínios funcionais de uma rede contínua. A forma ramificada do retículo permite contato com diferentes regiões do citoplasma, aproximação com outras organelas e participação em fluxos de lipídios, cálcio e proteínas. [Figura 6.8]
O complexo de Golgi, por sua vez, funciona como uma estação de processamento e distribuição. Proteínas e lipídios provenientes do retículo endoplasmático chegam a suas cisternas, passam por modificações químicas e são direcionados para destinos distintos. Em células animais secretoras, essa função é evidente na produção de hormônios, enzimas digestivas, anticorpos e componentes da matriz extracelular. Em células vegetais, o Golgi participa intensamente da produção e modificação de polissacarídeos da parede celular, além de contribuir para secreção, crescimento e remodelamento da superfície celular. A atividade do Golgi revela que a célula eucarionte não apenas sintetiza moléculas; ela as prepara, etiqueta, seleciona e envia ao local adequado. Essa lógica de processamento e endereçamento será detalhada posteriormente, mas já é suficiente reconhecer que a especialização das organelas está inseparavelmente ligada ao tráfego intracelular.
As vesículas fazem a comunicação operacional entre muitos desses compartimentos. Pequenas estruturas membranosas brotam de uma organela, carregam proteínas, lipídios ou solutos e se fundem com outra membrana de destino. Esse processo permite que o retículo endoplasmático envie material ao Golgi, que o Golgi distribua produtos para membrana plasmática, endossomos, lisossomos ou vacúolos, e que a membrana plasmática internalize componentes do meio externo por endocitose. O tráfego vesicular não é aleatório. Ele depende de sinais moleculares, proteínas de revestimento, proteínas de reconhecimento, pequenas GTPases e mecanismos de fusão que garantem especificidade. A célula eucarionte depende dessa precisão porque uma proteína enviada ao compartimento errado pode perder função, causar toxicidade ou comprometer a organização celular.
Lisossomos, vacúolos e endossomos mostram como a compartimentalização também serve à digestão, reciclagem e controle de qualidade. Em células animais, lisossomos concentram enzimas hidrolíticas capazes de degradar proteínas, lipídios, carboidratos e ácidos nucleicos em ambiente ácido. Eles recebem material por endocitose, fagocitose e autofagia, participando tanto da defesa celular quanto da renovação de componentes internos. Em células vegetais e fúngicas, vacúolos exercem funções parcialmente equivalentes, mas com amplitude ainda maior: armazenam íons, metabólitos, pigmentos, compostos de defesa e produtos de reserva, além de participarem da degradação e do controle osmótico. O vacúolo central de muitas células vegetais também contribui para o crescimento celular, pois sua expansão permite aumento de volume com menor custo de síntese citoplasmática. [Figura 6.9]
A relevância agrícola dessa organização é direta. A qualidade de frutos, sementes, folhas e raízes depende, em muitos casos, de processos vacuolares: acúmulo de açúcares, ácidos orgânicos, pigmentos, compostos fenólicos, proteínas de reserva, íons e moléculas associadas ao sabor, cor, defesa e valor nutricional. Em condições de salinidade, a capacidade de compartimentalizar Na⁺ no vacúolo pode reduzir a toxicidade citosólica e preservar enzimas sensíveis. Em déficit hídrico, o ajuste osmótico envolve solutos acumulados no citosol e no vacúolo, permitindo manutenção parcial do turgor e da atividade metabólica. Assim, uma organela frequentemente apresentada apenas como “armazenamento” participa de processos fisiológicos decisivos para produtividade, adaptação e qualidade pós-colheita. [Quadro 6.3]
Mitocôndrias e cloroplastos ampliam a discussão porque sua especialização funcional está ligada à conversão de energia. As mitocôndrias estão presentes na maioria das células eucarióticas e participam da respiração celular, integrando oxidação de combustíveis, ciclo do ácido cítrico, transporte de elétrons, formação de gradiente de prótons e síntese de ATP. Sua organização em membrana externa, espaço intermembrana, membrana interna e matriz permite separar etapas e criar condições eletroquímicas adequadas. A membrana interna, com suas cristas, amplia a superfície disponível para complexos proteicos envolvidos na fosforilação oxidativa. Essa organização revela um princípio recorrente: a forma da organela aumenta sua capacidade funcional porque cria área, compartimentos e gradientes.
Os cloroplastos, presentes em plantas e algas fotossintéticas, realizam outra forma de conversão energética. Suas membranas tilacoidais abrigam pigmentos, fotossistemas, cadeias transportadoras de elétrons e ATP sintase, enquanto o estroma contém enzimas envolvidas na fixação de carbono. Essa separação permite que a energia luminosa seja convertida em energia química e posteriormente utilizada na síntese de carboidratos. A especialização dos cloroplastos não se limita à fotossíntese; plastídios também participam da biossíntese de aminoácidos, lipídios, pigmentos e metabólitos especializados. Em células de raízes, frutos ou sementes, plastídios não fotossintéticos podem armazenar amido, pigmentos ou lipídios, mostrando que uma mesma linhagem de organelas pode assumir funções distintas conforme o tipo celular e o estágio de desenvolvimento.
A origem endossimbiótica de mitocôndrias e cloroplastos reforça a singularidade dessas organelas. Suas duplas membranas, seus genomas próprios, seus ribossomos e sua divisão por processos relacionados à fissão indicam descendência de bactérias ancestrais incorporadas por células precursoras e mantidas ao longo da evolução. No entanto, a autonomia atual dessas organelas é limitada. A maior parte de suas proteínas é codificada por genes nucleares, sintetizada em ribossomos citosólicos e importada posteriormente. Essa dependência cria uma integração profunda entre núcleo, citosol e organelas energéticas. A mitocôndria e o cloroplasto não são “células dentro da célula” em sentido pleno; são organelas semiautônomas cuja função depende da coordenação entre genomas, sinais e sistemas de transporte.
Os peroxissomos representam outro tipo de especialização. São organelas delimitadas por membrana simples, envolvidas em reações oxidativas, metabolismo de peróxido de hidrogênio, degradação de ácidos graxos e processos específicos em células animais e vegetais. A presença da catalase permite degradar H₂O₂, evitando que essa molécula se acumule em níveis prejudiciais. Em plantas, peroxissomos participam de processos associados à fotorrespiração, metabolismo lipídico durante a germinação de sementes oleaginosas e respostas a estresses oxidativos. Sua importância muitas vezes é subestimada porque sua morfologia parece simples em comparação com mitocôndrias ou cloroplastos, mas sua função metabólica é estratégica. A especialização de uma organela não depende de complexidade visual; depende da rede de reações que ela concentra e da integração que estabelece com o restante da célula.
O núcleo ocupa uma posição particular entre as organelas membranosas. Ele delimita o espaço onde o DNA eucariótico é armazenado, compactado, replicado, reparado e transcrito. O envoltório nuclear, formado por dupla membrana, separa o conteúdo nuclear do citosol, mas não isola completamente os dois ambientes. Complexos de poro nuclear controlam a passagem de proteínas, RNAs e complexos ribonucleoproteicos, permitindo comunicação intensa entre núcleo e citoplasma. Essa separação tem consequências profundas: a transcrição ocorre no núcleo, enquanto a tradução ocorre no citosol ou associada ao retículo endoplasmático. Entre uma etapa e outra, o RNA pode ser processado, selecionado, exportado ou degradado. Assim, a compartimentalização nuclear acrescenta camadas de regulação à expressão gênica. [Figura 6.10]
A existência de organelas membranosas também explica por que a célula eucarionte precisa de sistemas de endereçamento molecular. Uma proteína destinada ao núcleo precisa conter informações reconhecidas por receptores de importação nuclear. Uma proteína mitocondrial precisa apresentar sinais que permitam sua passagem por complexos de translocação nas membranas mitocondriais. Proteínas de cloroplastos, peroxissomos, retículo endoplasmático, lisossomos ou vacúolos também dependem de sinais específicos. Sem esses mecanismos, a compartimentalização perderia sentido, pois os componentes não chegariam ao lugar adequado. A célula eucarionte resolve esse desafio combinando síntese citosólica, sinais de localização, receptores, canais de translocação, vesículas e mecanismos de retenção. A identidade de cada organela depende não apenas do que ela contém, mas da capacidade de importar, exportar e preservar seus componentes corretos.
A descoberta experimental desses mecanismos foi uma das grandes conquistas da biologia celular moderna. O uso combinado de microscopia eletrônica, fracionamento celular, marcação radioativa de proteínas recém-sintetizadas, imunolocalização, proteínas fluorescentes e sistemas livres de células permitiu acompanhar o caminho de moléculas dentro da célula. Experimentos de pulso e perseguição, por exemplo, mostraram que proteínas secretadas podiam ser rastreadas desde o retículo endoplasmático até o Golgi e, posteriormente, até vesículas secretoras. Estudos sobre sequências-sinal demonstraram que a informação necessária ao destino de uma proteína podia estar contida em trechos específicos da própria cadeia polipeptídica. Esses resultados mudaram a interpretação da organela: ela deixou de ser apenas uma estrutura observada ao microscópio e passou a ser entendida como parte de uma rota molecular organizada.
As organelas membranosas não funcionam isoladamente. O retículo endoplasmático estabelece contatos com mitocôndrias, peroxissomos, membrana plasmática, endossomos e cloroplastos em células vegetais. Esses contatos não são fusões completas, mas aproximações especializadas que permitem troca de lipídios, sinalização por cálcio, coordenação metabólica e resposta a estresses. Mitocôndrias podem ajustar sua atividade conforme sinais citosólicos e necessidades energéticas. Cloroplastos comunicam seu estado funcional ao núcleo, modulando expressão gênica por sinalização retrógrada. Peroxissomos interagem metabolicamente com mitocôndrias e cloroplastos, especialmente em processos oxidativos e na fotorrespiração. O sistema de endomembranas conecta síntese, modificação, secreção, endocitose e degradação. A célula eucarionte funciona porque essas relações são coordenadas.
Em células vegetais, a integração entre organelas é especialmente evidente durante a fotossíntese e a fotorrespiração. O cloroplasto fixa carbono e produz compostos que alimentam o metabolismo celular, mas sua atividade gera demandas de equilíbrio redox, consumo de ATP, exportação de açúcares fosfatados e coordenação com mitocôndrias e peroxissomos. Em condições de alta luminosidade, seca ou limitação de CO₂, o risco de produção excessiva de espécies reativas aumenta, exigindo participação de sistemas antioxidantes distribuídos entre cloroplastos, peroxissomos, mitocôndrias e citosol. A produtividade de uma planta cultivada depende, em parte, da capacidade de manter essa integração sob variações ambientais. Por isso, compreender organelas membranosas não é apenas conhecer uma lista de nomes; é entender como a célula vegetal preserva metabolismo, crescimento e sobrevivência em ambientes instáveis.
A especialização funcional das organelas também sustenta a diferenciação celular. Células secretoras apresentam retículo endoplasmático rugoso e Golgi desenvolvidos; células musculares possuem muitas mitocôndrias e sistemas especializados de armazenamento de cálcio; células fagocíticas têm intensa atividade endocítica e lisossomal; células do mesófilo foliar possuem abundantes cloroplastos; células de sementes podem apresentar plastídios e vacúolos adaptados ao acúmulo de reservas. A morfologia interna da célula reflete suas demandas funcionais. Em outras palavras, a célula reorganiza seus compartimentos conforme o papel que desempenha no organismo. Essa plasticidade mostra que as organelas não são componentes estáticos herdados de um modelo único de célula, mas estruturas ajustáveis à fisiologia, ao desenvolvimento e ao ambiente.
Essa visão tem implicações diretas para biotecnologia. A produção de proteínas recombinantes, por exemplo, depende do destino intracelular da proteína produzida. Uma proteína destinada à secreção precisa entrar no retículo endoplasmático, dobrar-se corretamente, passar pelo Golgi e ser exportada. Uma enzima que exige modificações pós-traducionais específicas pode não funcionar se for produzida em um sistema celular inadequado. Em plantas transgênicas, a localização de uma proteína no citosol, cloroplasto, vacúolo ou apoplasto pode alterar estabilidade, rendimento, segurança e função. Biofábricas vegetais, melhoramento de qualidade nutricional, resistência a estresses e engenharia metabólica dependem da capacidade de direcionar moléculas aos compartimentos corretos. A compartimentalização, nesse contexto, deixa de ser apenas um princípio estrutural e torna-se ferramenta tecnológica.
A organização das organelas também impõe limites. A célula precisa gastar energia para manter gradientes, transportar proteínas, acidificar compartimentos, movimentar vesículas, reparar membranas e degradar componentes defeituosos. A complexidade eucariótica tem custo metabólico. Esse custo, contudo, é compensado pela capacidade de controle, especialização e adaptação. A célula pode responder a mudanças ambientais redistribuindo enzimas, alterando tráfego vesicular, modificando a morfologia mitocondrial, ajustando a posição dos cloroplastos, expandindo o vacúolo, ativando autofagia ou reorganizando o retículo endoplasmático. Em condições de estresse severo, a célula pode degradar organelas danificadas e reciclar seus componentes. A especialização funcional, portanto, não significa rigidez; significa capacidade de reorganização controlada.
Um erro didático recorrente é apresentar organelas como se cada uma tivesse uma única função: mitocôndria produz ATP, cloroplasto faz fotossíntese, Golgi empacota proteínas, lisossomo digere, vacúolo armazena, núcleo guarda DNA. Essas frases ajudam na iniciação, mas empobrecem a compreensão. Mitocôndrias participam também de metabolismo intermediário, sinalização e morte celular. Cloroplastos produzem metabólitos, integram sinais ambientais e comunicam seu estado ao núcleo. O Golgi participa da biossíntese de componentes da parede celular em plantas. Vacúolos regulam osmose, degradação, armazenamento, defesa e crescimento. Peroxissomos integram metabolismo oxidativo, lipídico e respostas ao estresse. O núcleo não apenas guarda DNA, mas organiza cromatina, regula transcrição, processa RNAs e coordena programas celulares. A especialização das organelas deve ser entendida como predominância funcional, não como exclusividade.
A célula eucarionte, vista por esse ângulo, é uma rede de compartimentos especializados que trocam matéria, energia e informação. O citosol conecta, as membranas delimitam, as vesículas transportam, os sinais direcionam e as organelas executam etapas específicas de processos maiores. A função celular emerge dessa combinação. Nenhuma organela possui sentido completo fora da célula que a integra, e nenhuma célula eucarionte mantém sua organização sem a identidade funcional de suas organelas. A compartimentalização cria a possibilidade da especialização, mas a integração impede que a especialização se transforme em fragmentação. Essa é uma das chaves para compreender os capítulos seguintes: cada organela poderá ser estudada separadamente por necessidade analítica, mas a célula viva opera como uma unidade coordenada, dinâmica e profundamente interdependente.
6.4 — Sistema de endomembranas: visão geral da rota secretora e endocítica #
A célula eucarionte não apenas possui organelas membranosas; ela mantém entre muitas delas um sistema contínuo de comunicação, renovação e distribuição de componentes. Essa rede, conhecida como sistema de endomembranas, reúne compartimentos que trocam membranas, proteínas, lipídios e solutos por meio de conexões diretas ou de vesículas de transporte. O envoltório nuclear, o retículo endoplasmático, o complexo de Golgi, endossomos, lisossomos, vacúolos, vesículas secretoras e a própria membrana plasmática participam dessa organização. A célula, nesse sentido, não produz moléculas para deixá-las dispersas ao acaso. Ela sintetiza, modifica, seleciona, empacota, transporta, secreta, internaliza, recicla e degrada componentes por rotas controladas, mantendo a composição de suas membranas e a identidade funcional de seus compartimentos. [Figura 6.11]
A lógica do sistema de endomembranas começa com uma propriedade essencial das membranas biológicas: elas podem se deformar, brotar, fundir-se e conservar sua orientação. Quando uma vesícula se forma a partir de uma membrana doadora, a face citosólica da membrana permanece voltada para o citosol, enquanto a face luminal preserva equivalência topológica com o interior dos compartimentos ou com o espaço extracelular após fusão com a membrana plasmática. Essa continuidade topológica permite compreender por que proteínas destinadas à secreção, ao lúmen de organelas ou à superfície celular seguem rotas específicas desde sua síntese. A célula eucarionte utiliza membranas não apenas como barreiras, mas como superfícies móveis capazes de preservar relações de orientação, composição e destino. [Figura 6.12]
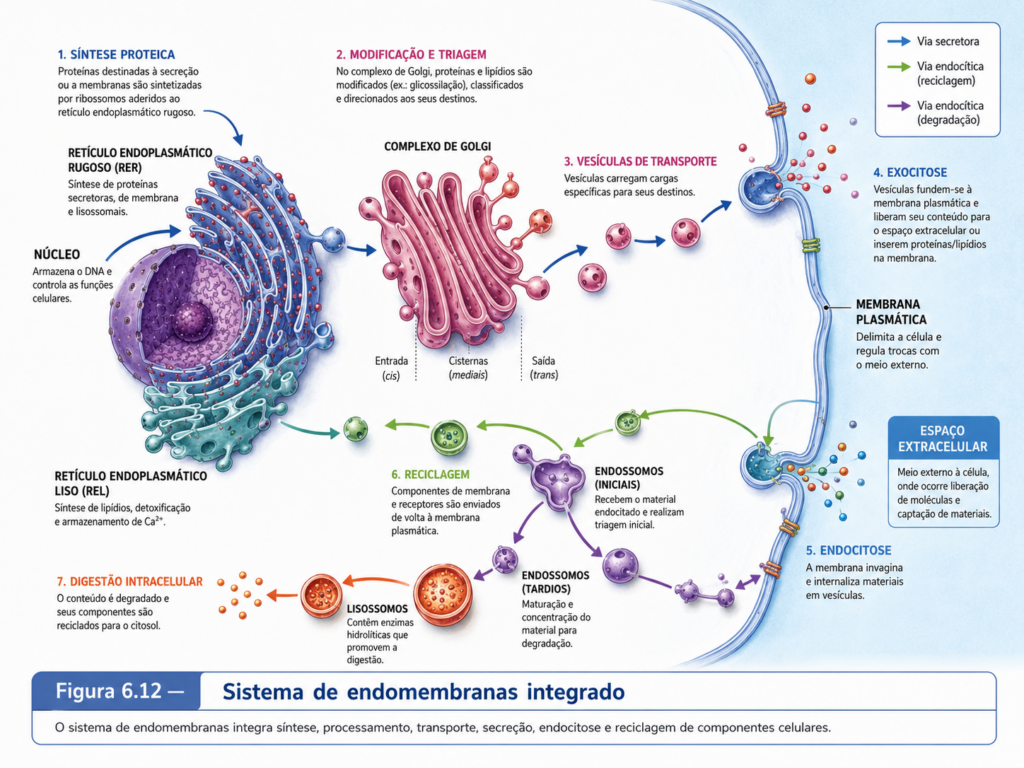
O retículo endoplasmático ocupa posição inicial decisiva nesse sistema. Sua membrana é contínua com a membrana externa do envoltório nuclear, formando uma rede extensa que se distribui pelo citoplasma. Nas regiões associadas a ribossomos, o retículo endoplasmático rugoso recebe proteínas cuja síntese é direcionada para a rota secretora. Essas proteínas podem ser translocadas para o lúmen do retículo ou inseridas em sua membrana à medida que são sintetizadas. No lúmen, muitas delas iniciam processos de dobramento, formação de pontes dissulfeto, montagem em complexos e modificações químicas iniciais, como glicosilação. A entrada no retículo não é um evento aleatório; depende de sinais presentes na cadeia polipeptídica nascente, reconhecidos por mecanismos que conduzem o ribossomo à membrana do retículo. Essa etapa transforma informação contida na sequência da proteína em localização intracelular. [Animação 6.2]
O retículo endoplasmático liso, embora não apresente ribossomos aderidos em sua superfície, também participa da organização do sistema. Ele atua na síntese de lipídios, no metabolismo de moléculas hidrofóbicas e, em determinados tipos celulares, no armazenamento e liberação de cálcio. A produção de lipídios no retículo é particularmente importante porque novas membranas precisam ser continuamente formadas, expandidas, remodeladas e distribuídas. A célula não herda suas membranas como estruturas fixas; ela as reconstrói continuamente. A membrana plasmática, as vesículas, o Golgi, os endossomos e outros compartimentos dependem, em grande parte, da síntese e redistribuição de lipídios iniciada no retículo endoplasmático. Essa renovação torna o sistema de endomembranas uma rede dinâmica, não um conjunto estático de compartimentos.
A saída de proteínas e lipídios do retículo endoplasmático ocorre por vesículas que seguem em direção ao complexo de Golgi. O Golgi funciona como uma estação de processamento, triagem e redistribuição, composta por cisternas organizadas em regiões funcionalmente distintas. Moléculas recém-chegadas podem sofrer modificações adicionais, especialmente em cadeias de carboidratos associadas a proteínas e lipídios. Essas modificações não são enfeites moleculares. Elas podem influenciar estabilidade, reconhecimento, atividade, destino e interação celular. Em células animais, o Golgi participa da produção de glicoproteínas, proteoglicanos e componentes da matriz extracelular. Em células vegetais, sua relevância é ainda mais evidente na produção de polissacarídeos não celulósicos da parede celular, como pectinas e hemiceluloses, que serão enviados à superfície celular por vesículas secretoras. [Tabela 6.4]
A passagem pelo Golgi revela um princípio central da rota secretora: a célula precisa distinguir destinos. Algumas moléculas serão secretadas para o meio extracelular; outras permanecerão na membrana plasmática; outras seguirão para endossomos, lisossomos ou vacúolos; outras retornarão a compartimentos anteriores. Essa seleção exige sinais moleculares e mecanismos de reconhecimento. Vesículas diferentes carregam conjuntos específicos de proteínas de revestimento, adaptadores, receptores de carga e marcadores de destino. Pequenas GTPases, proteínas de ancoragem e proteínas de fusão garantem que uma vesícula se aproxime e se funda com a membrana correta. Esse nível de especificidade é indispensável porque a mistura indevida de componentes comprometeria a identidade dos compartimentos. Um lisossomo não pode perder sua composição hidrolítica e ácida; a membrana plasmática não pode receber indiscriminadamente proteínas destinadas a compartimentos internos; o retículo precisa recuperar proteínas residentes que escapam para o Golgi.
A secreção representa uma das faces mais evidentes desse sistema. Em muitas células, vesículas derivadas do Golgi transportam proteínas, lipídios e carboidratos até a membrana plasmática, onde se fundem e liberam seu conteúdo para o exterior. Esse processo sustenta funções tão diversas quanto liberação de hormônios, secreção de enzimas digestivas, produção de anticorpos, deposição de matriz extracelular, formação da parede celular vegetal e renovação da superfície celular. A exocitose também acrescenta componentes à membrana plasmática, pois a membrana da vesícula passa a integrar a membrana celular após a fusão. Assim, secreção e expansão de membrana são processos conectados. A célula não apenas elimina produtos; ela remodela sua interface com o ambiente. [Figura 6.13]
Nas plantas, a rota secretora possui papel estrutural decisivo. Componentes da parede celular são produzidos ou processados em diferentes compartimentos e enviados à superfície por vesículas. A expansão celular, a formação de tecidos, a resistência mecânica, a adesão entre células e as respostas a patógenos dependem da produção e entrega coordenada desses materiais. Durante o crescimento de uma célula vegetal, a parede precisa ser suficientemente rígida para sustentar a forma e suficientemente remodelável para permitir expansão. Essa aparente contradição é resolvida por um fluxo regulado de enzimas, polissacarídeos, proteínas de parede e componentes de membrana. A rota secretora, nesse contexto, não é apenas um mecanismo de exportação; é um processo que constrói a arquitetura do corpo vegetal a partir da escala celular. [Quadro 6.4]
A endocitose representa o movimento inverso: a célula internaliza regiões da membrana plasmática e componentes do meio externo, formando vesículas que entram no sistema endomembranoso. Essa internalização pode servir à captura de nutrientes, à remoção de receptores da superfície, à reciclagem de proteínas de membrana, à modulação de sinais, à defesa celular ou à renovação da própria membrana. Em células animais, a endocitose mediada por receptores permite concentrar moléculas específicas antes da internalização, aumentando a eficiência de captação. Em células vegetais, a endocitose contribui para remodelar a membrana plasmática, regular transportadores, ajustar respostas hormonais e controlar a polaridade celular. A membrana plasmática, portanto, não é uma fronteira estável e imutável; ela está em fluxo contínuo com compartimentos internos. [Animação 6.3]
Após a internalização, vesículas endocíticas geralmente se fundem com endossomos, compartimentos que funcionam como estações de triagem. Alguns componentes são reciclados de volta à membrana plasmática; outros seguem para degradação em lisossomos ou vacúolos; outros ainda podem ser redirecionados para compartimentos específicos. Essa triagem é fundamental para a regulação celular. Um receptor de membrana que permanece indefinidamente ativo pode gerar sinalização excessiva; ao ser internalizado, pode ser degradado ou reciclado, modulando a intensidade e duração da resposta. Um transportador de íons pode ser removido da membrana em determinada condição ambiental e reinserido em outra. Uma proteína de adesão pode ser redistribuída durante migração celular, crescimento ou diferenciação. A endocitose, assim, permite que a célula ajuste sua superfície ao ambiente e ao seu estado fisiológico.
Lisossomos e vacúolos ocupam posição final em muitas rotas de degradação. Em células animais, os lisossomos recebem material vindo da endocitose, fagocitose e autofagia, degradando macromoléculas por enzimas hidrolíticas ativas em pH ácido. Em células vegetais e fúngicas, vacúolos desempenham funções degradativas e de armazenamento, combinando digestão, reciclagem, controle osmótico e acúmulo de substâncias. A degradação não deve ser vista apenas como destruição. Ela é parte da economia celular. Ao degradar proteínas, organelas danificadas, patógenos ou material extracelular internalizado, a célula recupera aminoácidos, açúcares, nucleotídeos, lipídios e íons que podem ser reutilizados. Em períodos de escassez nutricional ou estresse, essa capacidade de reciclagem pode ser decisiva para a sobrevivência celular.
A autofagia amplia essa lógica ao permitir que componentes internos sejam envoltos por membranas e encaminhados à degradação. Mitocôndrias danificadas, porções do retículo endoplasmático, agregados proteicos e regiões do citoplasma podem ser seletivamente removidos. Esse processo mantém a qualidade interna da célula e permite adaptação a mudanças nutricionais, envelhecimento, estresse oxidativo e danos celulares. Em plantas, a autofagia participa da remobilização de nutrientes, da senescência foliar, da resposta a estresses e da defesa contra patógenos. Uma folha em senescência, por exemplo, não simplesmente “morre”; ela remobiliza nitrogênio, carbono e outros recursos para órgãos em crescimento, sementes ou estruturas de reserva. Essa remobilização depende de sistemas degradativos coordenados, nos quais vacúolos e tráfego de membranas exercem papel central.
A rota secretora e a rota endocítica não são sistemas independentes. Elas se equilibram continuamente. A exocitose adiciona membrana à superfície celular; a endocitose remove membrana. A secreção entrega proteínas e carboidratos ao exterior; a internalização recupera receptores, transportadores e moléculas do ambiente. O Golgi envia componentes adiante; rotas retrógradas devolvem proteínas ao retículo ou a compartimentos anteriores. Endossomos selecionam entre reciclagem e degradação. Lisossomos e vacúolos recebem material de várias origens. Essa circulação mantém a composição das membranas, ajusta o tamanho dos compartimentos e permite respostas rápidas a mudanças externas. A célula eucarionte preserva sua identidade não por imobilidade, mas por fluxo controlado.
A manutenção da identidade de cada compartimento depende de mecanismos moleculares precisos. Compartimentos do sistema de endomembranas possuem conjuntos próprios de lipídios, proteínas residentes, enzimas, receptores, marcadores moleculares e condições internas. Mesmo quando há troca constante de vesículas, cada organela conserva características específicas. Essa identidade é mantida por sinais de retenção, recuperação seletiva, direcionamento vesicular, maturação progressiva de compartimentos e atividade de bombas iônicas. A acidificação de endossomos, lisossomos e vacúolos, por exemplo, não é consequência passiva do isolamento; ela depende de bombas de prótons que consomem energia para criar um ambiente interno adequado à função degradativa e à triagem molecular.
O sistema de endomembranas também está ligado ao controle de qualidade das proteínas. No retículo endoplasmático, proteínas destinadas à secreção ou à membrana precisam dobrar-se corretamente antes de seguir adiante. Chaperonas e enzimas residentes auxiliam esse processo. Proteínas mal dobradas podem ser retidas, redirecionadas para degradação ou desencadear respostas celulares quando se acumulam em excesso. Essa vigilância é essencial porque proteínas secretadas ou de membrana frequentemente atuam em comunicação, transporte, adesão, defesa e reconhecimento. Um erro de dobramento ou endereçamento pode comprometer funções celulares e, em organismos multicelulares, produzir doenças ou defeitos de desenvolvimento. A célula, portanto, avalia a qualidade de parte de sua produção antes de permitir que ela avance pela rota secretora.
A história experimental desse sistema mostra como a biologia celular deixou de ser apenas descritiva. A microscopia eletrônica permitiu visualizar retículo endoplasmático, Golgi, vesículas e lisossomos com riqueza estrutural, mas a compreensão do fluxo entre esses compartimentos exigiu abordagens funcionais. Experimentos de pulso e perseguição com aminoácidos radioativos permitiram acompanhar proteínas recém-sintetizadas ao longo da rota secretora, mostrando sua passagem sequencial pelo retículo, Golgi e vesículas secretoras. Posteriormente, sistemas genéticos em leveduras revelaram genes necessários à secreção e ao tráfego vesicular, enquanto ensaios bioquímicos identificaram proteínas de revestimento, GTPases e mecanismos de fusão. A ideia moderna de sistema de endomembranas nasceu dessa convergência entre imagem, rastreamento molecular, genética e bioquímica.
A levedura Saccharomyces cerevisiae tornou-se particularmente importante nesse campo porque combina organização eucariótica com facilidade de manipulação genética. Mutantes com defeitos em secreção permitiram identificar etapas específicas do tráfego intracelular. Quando uma proteína necessária à formação ou fusão de vesículas era alterada, moléculas destinadas à secreção acumulavam-se em compartimentos específicos, revelando a ordem dos eventos. Esse tipo de experimento mostra uma lição metodológica importante: para compreender uma rota celular, muitas vezes é necessário interrompê-la em pontos definidos e observar onde o fluxo se acumula. A célula viva é dinâmica demais para ser entendida apenas por imagens estáticas; suas rotas precisam ser rastreadas no tempo. [Figura 6.14]
O tráfego vesicular tem forte importância biotecnológica. A produção de proteínas recombinantes, vacinas, enzimas industriais, anticorpos, hormônios e moléculas terapêuticas depende do sistema celular usado como plataforma. Uma proteína que precisa ser secretada deve entrar corretamente no retículo endoplasmático, dobrar-se, passar pelo Golgi e ser liberada sem degradação. Uma proteína que requer glicosilação específica pode ter propriedades diferentes se for produzida em bactérias, leveduras, células animais ou plantas. Em biofábricas vegetais, a decisão de direcionar uma proteína ao apoplasto, ao vacúolo, ao retículo endoplasmático ou ao cloroplasto pode alterar rendimento, estabilidade, atividade e segurança. Assim, a rota secretora não é apenas um tema de biologia celular básica; ela é um eixo operacional da biotecnologia moderna.
Nas ciências agrárias, o sistema de endomembranas aparece em processos que vão da germinação à defesa vegetal. Durante a germinação de sementes, reservas armazenadas precisam ser degradadas e mobilizadas para sustentar o crescimento inicial da plântula. Durante a formação de parede celular, vesículas entregam polissacarídeos e proteínas à superfície. Na defesa contra patógenos, a secreção direcionada pode levar enzimas, compostos antimicrobianos e proteínas relacionadas à resposta imune para regiões de contato com o invasor. Na adaptação à salinidade, transportadores podem ser redistribuídos entre membrana plasmática, endossomos e vacúolos, ajustando fluxos iônicos. Em cada caso, o tráfego interno de membranas participa da capacidade da planta de crescer, defender-se e adaptar-se.
A endocitose também possui papel regulatório nas respostas hormonais vegetais. A distribuição polar de transportadores de auxina, por exemplo, depende de processos de inserção, remoção e reciclagem de proteínas de membrana. Como a auxina influencia divisão, expansão, diferenciação e formação de órgãos, o tráfego endomembranoso participa indiretamente da arquitetura da planta. Uma alteração na localização de transportadores pode mudar gradientes hormonais, e uma mudança em gradientes hormonais pode modificar padrões de crescimento. Essa relação mostra que o sistema de endomembranas não atua apenas em funções domésticas de transporte; ele participa da construção de formas biológicas.
A membrana plasmática ocupa uma posição especial nesse sistema porque é, ao mesmo tempo, fronteira externa e ponto de continuidade com rotas internas. Receptores presentes na membrana detectam sinais do ambiente; canais e transportadores controlam entrada e saída de solutos; proteínas de adesão e parede celular participam de interações com o exterior; vesículas secretoras adicionam componentes à superfície; vesículas endocíticas removem componentes da superfície. A membrana plasmática é, portanto, uma interface em fluxo. Seu funcionamento depende tanto do ambiente externo quanto do sistema interno que a renova e reorganiza. A célula eucarionte não apenas responde ao meio; ela remodela ativamente a superfície por onde essa resposta começa.
A separação entre rota secretora e rota endocítica é útil para organizar o estudo, mas a célula opera com circuitos interligados. Uma proteína pode sair do retículo, passar pelo Golgi, alcançar a membrana plasmática, ser internalizada por endocitose, reciclada para a superfície ou enviada à degradação. Um receptor pode alternar entre presença superficial, internalização e reciclagem conforme o sinal recebido. Um transportador pode mudar de localização conforme disponibilidade de nutrientes, salinidade, pH ou estado energético. Essa plasticidade permite que a célula ajuste sua fisiologia sem precisar sintetizar tudo novamente a cada mudança. Redistribuir componentes existentes pode ser mais rápido e energeticamente eficiente do que produzir novos componentes do zero.
A dinâmica do sistema de endomembranas também depende do citoesqueleto. Vesículas precisam se deslocar por distâncias que, em escala molecular, podem ser grandes. Microtúbulos e filamentos de actina fornecem trilhos para proteínas motoras transportarem vesículas e organelas. Em células vegetais, a organização do tráfego vesicular está intimamente relacionada à expansão celular, à deposição de parede e à orientação do crescimento. Em células animais polarizadas, como neurônios e células epiteliais, o direcionamento vesicular garante que proteínas sejam entregues a regiões específicas da superfície. O tráfego de membranas, portanto, depende de uma colaboração contínua entre organelas, vesículas, citoesqueleto, motores moleculares e sinais de destino.
A organização desse sistema ajuda a explicar por que a célula eucarionte suporta maior complexidade funcional do que uma célula sem compartimentos internos membranosos equivalentes. Ao separar síntese, processamento, secreção, internalização e degradação, a célula cria pontos de controle sucessivos. Uma molécula pode ser avaliada no retículo, modificada no Golgi, selecionada em endossomos, reciclada ou degradada. Cada etapa oferece oportunidade de regulação. Essa multiplicidade de pontos de controle aumenta a precisão, mas também exige coordenação. Falhas no tráfego vesicular podem causar acúmulo de proteínas, defeitos de secreção, alteração da superfície celular, comprometimento da degradação lisossomal ou vacuolar e distúrbios na sinalização.
A célula eucarionte também usa o sistema de endomembranas para lidar com mudança e dano. Quando há estresse no retículo endoplasmático, respostas celulares tentam reduzir a carga de proteínas mal dobradas, aumentar a capacidade de dobramento ou degradar componentes problemáticos. Quando organelas são danificadas, processos autofágicos podem removê-las. Quando a superfície celular precisa ser remodelada, a endocitose e a reciclagem ajustam sua composição. Quando uma célula vegetal cresce em determinada direção, vesículas entregam componentes de membrana e parede em regiões específicas. A rede endomembranosa dá à célula capacidade de manutenção, adaptação e reconstrução.
No conjunto, o sistema de endomembranas representa uma infraestrutura interna de fluxo. Ele integra síntese, modificação, transporte, secreção, endocitose, reciclagem e degradação em uma rede que preserva a identidade dos compartimentos e permite remodelamento contínuo. Sua importância está menos na lista de estruturas envolvidas e mais na lógica que estabelece: a célula eucarionte mantém ordem por meio de circulação controlada. Moléculas e membranas se movem, mas não se dispersam; compartimentos trocam componentes, mas não perdem identidade; a superfície celular se renova, mas conserva seletividade. Essa combinação de movimento e controle é uma das bases da organização eucarionte e prepara a análise detalhada do retículo endoplasmático, do Golgi, do tráfego vesicular, da endocitose, da exocitose e da secreção nos capítulos seguintes.
6.5 — Organelas semiautônomas: mitocôndrias e cloroplastos na lógica eucarionte #
Entre as organelas membranosas da célula eucarionte, mitocôndrias e cloroplastos ocupam uma posição singular. Elas não são apenas compartimentos especializados delimitados por membranas, como ocorre com retículo endoplasmático, complexo de Golgi, lisossomos ou vacúolos. Mitocôndrias e cloroplastos conservam marcas profundas de uma história evolutiva própria, associada à incorporação de organismos procarióticos ancestrais por células precursoras dos eucariontes modernos. Essa origem endossimbiótica ajuda a explicar por que essas organelas apresentam dupla membrana, DNA próprio, ribossomos próprios e capacidade de divisão por processos semelhantes à fissão. Ao mesmo tempo, sua autonomia atual é limitada, porque a maior parte de suas proteínas é codificada por genes nucleares, sintetizada no citosol e posteriormente importada para o interior da organela. Por isso, a expressão “semiautônomas” é mais precisa do que “autônomas”: mitocôndrias e cloroplastos mantêm traços de independência evolutiva, mas funcionam hoje como partes integradas de uma célula eucarionte regulada pelo núcleo, pelo citosol e por redes metabólicas compartilhadas. [Figura 6.15]
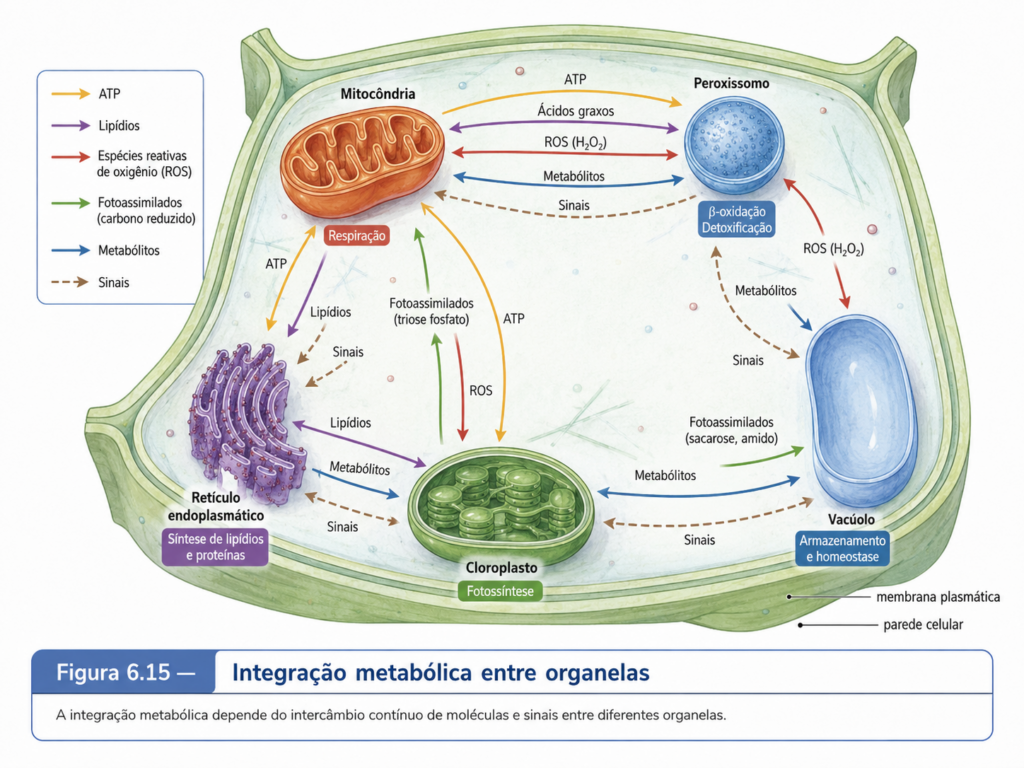
A teoria endossimbiótica propõe que as mitocôndrias derivam de bactérias aeróbias ancestrais incorporadas por uma célula hospedeira, enquanto os cloroplastos derivam de cianobactérias fotossintéticas incorporadas por ancestrais de algas e plantas. Essa interpretação não se baseia apenas em semelhanças superficiais. A presença de DNA circular, ribossomos semelhantes aos bacterianos, divisão por fissão, dupla membrana e relações filogenéticas entre genes organelares e genes bacterianos constitui um conjunto robusto de evidências. A membrana interna dessas organelas carrega muitas características funcionais relacionadas ao antigo procarionte incorporado, enquanto a membrana externa pode ser interpretada como parte da interface estabelecida durante o processo de internalização e integração celular. Essa origem explica por que mitocôndrias e cloroplastos não se encaixam completamente na lógica do sistema de endomembranas, embora mantenham comunicação intensa com ele e com o restante da célula. [Figura 6.16]
O estabelecimento dessas endossimbioses foi uma transição decisiva na evolução celular. A incorporação de uma bactéria aeróbia capaz de utilizar oxigênio para extrair energia de moléculas orgânicas teria ampliado enormemente a eficiência energética da célula hospedeira. Em um planeta cuja atmosfera passou a acumular oxigênio em consequência da atividade fotossintética de organismos procarióticos, a capacidade de usar esse gás como aceptor final de elétrons tornou-se uma vantagem metabólica poderosa. O oxigênio, que podia ser tóxico para formas celulares não adaptadas, passou a ser explorado como parte de uma maquinaria eficiente de produção de ATP. A mitocôndria representa, nesse sentido, uma solução evolutiva para o aproveitamento controlado do metabolismo aeróbio dentro de uma célula maior e mais complexa.
A origem dos cloroplastos acrescentou outra dimensão à história eucariótica. Ao incorporar uma célula fotossintética ancestral, a linhagem que daria origem a algas e plantas adquiriu a capacidade de converter energia luminosa em energia química, fixar carbono e sustentar cadeias tróficas inteiras. A fotossíntese eucariótica não surgiu como simples invenção interna de uma célula preexistente, mas como integração progressiva de uma maquinaria procariótica ao contexto celular eucarionte. Os cloroplastos conservaram tilacoides, pigmentos, fotossistemas e mecanismos de transporte de elétrons relacionados à sua origem cianobacteriana, mas passaram a depender da célula hospedeira para a maior parte de sua biogênese, divisão, manutenção e regulação. A planta moderna, portanto, é inseparável dessa dupla herança: eucariótica na organização nuclear e citoplasmática, procariótica na origem de seus plastídios fotossintéticos. [Tabela 6.5]
Apesar da presença de DNA próprio, os genomas mitocondriais e cloroplastidiais atuais são muito menores do que os genomas das bactérias das quais essas organelas descendem. Ao longo da evolução, muitos genes foram perdidos ou transferidos para o núcleo. Essa transferência gênica teve consequências profundas. Proteínas que antes eram codificadas e produzidas dentro do organismo endossimbionte passaram a ser codificadas no genoma nuclear, sintetizadas por ribossomos citosólicos e importadas de volta para a organela. Essa reorganização criou uma dependência recíproca: a organela preservou funções metabólicas essenciais, enquanto o núcleo assumiu grande parte do controle genético sobre sua composição proteica. A célula eucarionte tornou-se, assim, um sistema genético distribuído, no qual diferentes genomas precisam funcionar de maneira coordenada.
A importação de proteínas é uma das chaves dessa coordenação. A maioria das proteínas mitocondriais é sintetizada no citosol como precursores contendo sinais de endereçamento que permitem seu reconhecimento por complexos de translocação localizados nas membranas mitocondriais. Após atravessar a membrana externa e, em muitos casos, também a membrana interna, essas proteínas alcançam matriz, espaço intermembrana, membrana interna ou membrana externa, conforme suas informações de destino. Nos cloroplastos ocorre processo semelhante, embora com especificidades próprias: proteínas codificadas no núcleo são sintetizadas no citosol, reconhecidas por sequências de trânsito e importadas através de complexos localizados nas membranas externa e interna do envelope plastidial. Algumas ainda precisam ser direcionadas posteriormente aos tilacoides, ao estroma, ao envelope ou a outros domínios internos. O destino final de uma proteína depende de sinais moleculares que funcionam como endereços intracelulares. [Figura 6.17]
Esse sistema de importação mostra que a autonomia das organelas é parcial. Mitocôndrias e cloroplastos produzem algumas de suas próprias proteínas, mas não possuem sozinhos todos os elementos necessários para sua formação e funcionamento. Seus ribossomos sintetizam um conjunto limitado de polipeptídios, geralmente componentes centrais de complexos envolvidos em transporte de elétrons e conversão de energia. Entretanto, esses complexos também dependem de numerosas subunidades codificadas no núcleo. A montagem de uma cadeia respiratória mitocondrial ou de um fotossistema cloroplastidial exige, portanto, coordenação precisa entre síntese citosólica, importação organelar, expressão do genoma da organela, montagem de complexos proteicos e inserção em membranas internas. A função energética depende de cooperação entre compartimentos genéticos e celulares.
A mitocôndria ilustra essa integração de maneira particularmente clara. Ela participa da oxidação de combustíveis orgânicos, do ciclo do ácido cítrico, da cadeia transportadora de elétrons, da fosforilação oxidativa, do metabolismo de aminoácidos e lipídios, do controle redox, da sinalização celular e, em muitos eucariontes, de processos associados à morte celular programada. Sua arquitetura interna reflete essa multiplicidade. A membrana externa permite comunicação seletiva com o citosol; o espaço intermembrana participa de gradientes e sinalização; a membrana interna abriga os complexos respiratórios e a ATP sintase; a matriz contém enzimas metabólicas, DNA mitocondrial, ribossomos e sistemas próprios de expressão gênica. As cristas mitocondriais ampliam a área da membrana interna e organizam espacialmente a maquinaria de produção de ATP. A forma da organela, nesse caso, está diretamente relacionada à intensidade e à regulação de sua função.
A mitocôndria, entretanto, não deve ser reduzida à expressão simplificada de “usina de energia da célula”. Essa analogia popular é útil apenas até certo ponto. A mitocôndria não apenas produz ATP; ela integra metabolismo, disponibilidade de nutrientes, estado redox, biossíntese, sinalização por cálcio, produção controlada de espécies reativas de oxigênio e respostas a danos celulares. Em tecidos com alta demanda energética, como músculos, neurônios, células secretoras e meristemas ativos, a quantidade, morfologia e distribuição das mitocôndrias acompanham as necessidades fisiológicas da célula. Em plantas, mitocôndrias precisam funcionar em diálogo com cloroplastos, peroxissomos e citosol, especialmente porque a fotossíntese, a respiração e a fotorrespiração criam fluxos metabólicos interdependentes. [Figura 6.18]
Os cloroplastos também não podem ser reduzidos a uma fórmula simplificada. Embora sejam as organelas centrais da fotossíntese em plantas e algas, eles participam de uma rede metabólica muito mais ampla. Além da captura de luz e fixação de carbono, plastídios contribuem para a síntese de aminoácidos, ácidos graxos, pigmentos, hormônios e metabólitos especializados. Em células fotossintéticas, os cloroplastos apresentam envelope duplo, estroma e sistema de tilacoides, onde se localizam os fotossistemas e outros componentes das reações dependentes de luz. No estroma, ocorrem reações relacionadas à assimilação de carbono e à biossíntese de diversas moléculas. Em tecidos não fotossintéticos, plastídios podem diferenciar-se em amiloplastos, cromoplastos, etioplastos ou outros tipos, especializados em armazenamento de amido, acúmulo de pigmentos, desenvolvimento em ausência de luz ou metabolismo específico de determinado tecido. A plasticidade dos plastídios é uma das bases celulares da diversidade funcional das plantas.
Essa plasticidade tem implicações agronômicas diretas. A cor de frutos e flores depende, em muitos casos, da diferenciação de plastídios e do acúmulo de carotenoides ou outros pigmentos. O armazenamento de amido em raízes, tubérculos, sementes e grãos envolve amiloplastos. A eficiência fotossintética de folhas depende da quantidade, distribuição e funcionamento dos cloroplastos. A resposta a estresses ambientais exige ajuste da atividade cloroplastidial, controle de excesso de energia luminosa, proteção contra espécies reativas de oxigênio e comunicação com o núcleo. Em culturas agrícolas submetidas a calor, seca, salinidade ou alta irradiância, os cloroplastos podem deixar de ser apenas locais de produção de açúcares e tornar-se fontes de sinais de estresse que reprogramam a fisiologia celular. [Quadro 6.5]
A comunicação entre cloroplastos e núcleo é essencial para a sobrevivência vegetal. Como grande parte das proteínas cloroplastidiais é codificada no núcleo, o desenvolvimento e a manutenção dos cloroplastos dependem da expressão coordenada de genes nucleares e plastidiais. Sinais originados no cloroplasto podem influenciar a expressão gênica nuclear, processo frequentemente chamado de sinalização retrógrada. Esses sinais podem refletir o estado redox, a disponibilidade de intermediários metabólicos, a integridade dos fotossistemas, a produção de espécies reativas de oxigênio ou o estágio de desenvolvimento plastidial. A célula vegetal precisa saber se seus cloroplastos estão funcionando adequadamente para ajustar síntese de proteínas fotossintéticas, mecanismos antioxidantes, metabolismo e crescimento. O cloroplasto, portanto, não é apenas executor de instruções nucleares; ele também informa ao núcleo seu estado funcional.
A mitocôndria também participa de comunicação retrógrada. Alterações na atividade respiratória, no potencial de membrana, no estado redox, na produção de espécies reativas de oxigênio ou na disponibilidade de metabólitos podem afetar a expressão de genes nucleares. Em plantas, essa comunicação se torna ainda mais complexa porque mitocôndrias e cloroplastos coexistem em uma mesma célula e compartilham demandas energéticas, metabólicas e redox. Durante o dia, cloroplastos produzem açúcares e energia química; mitocôndrias continuam atuando no metabolismo respiratório e na integração de intermediários; peroxissomos participam da fotorrespiração; o citosol conecta fluxos de carbono, nitrogênio e energia. A célula vegetal opera como uma rede metabólica multiorganelar, e mitocôndrias e cloroplastos são dois de seus centros mais importantes.
A presença de genomas próprios em mitocôndrias e cloroplastos impõe desafios particulares. Como cada célula pode conter muitas cópias dessas organelas e muitos genomas organelares, a herança e a expressão desses DNA não seguem exatamente os mesmos princípios da genética nuclear. Em muitos organismos, a herança mitocondrial ocorre predominantemente pela linhagem materna, embora existam variações. Em plantas, os genomas de organelas podem apresentar tamanhos, organizações e padrões de recombinação complexos. Além disso, mutações em DNA mitocondrial ou cloroplastidial podem afetar apenas parte das cópias presentes em uma célula ou tecido, criando situações em que genomas normais e mutantes coexistem. Essa condição, conhecida como heteroplasmia em contextos mitocondriais, mostra que a genética celular eucariótica envolve níveis adicionais de complexidade além do genoma nuclear.
Em plantas cultivadas, a genética de organelas possui relevância prática. Características relacionadas à macho-esterilidade citoplasmática, por exemplo, frequentemente envolvem interações entre genomas mitocondriais e nucleares e são exploradas em programas de produção de híbridos. A transformação de cloroplastos, embora tecnicamente mais restrita a algumas espécies e sistemas, tem interesse biotecnológico porque pode permitir alta expressão de proteínas, contenção parcial de transgenes pela herança plastidial em determinadas situações e engenharia de vias metabólicas localizadas no plastídio. A compreensão da semiautonomia das organelas, nesse contexto, não é apenas uma questão evolutiva; ela influencia melhoramento, biotecnologia, produção de biomoléculas e estratégias de manejo genético.
A divisão de mitocôndrias e cloroplastos também revela sua origem peculiar. Essas organelas não surgem de novo a partir do citosol; elas se multiplicam a partir de organelas preexistentes. Durante o crescimento celular e antes da divisão celular, mitocôndrias e plastídios precisam aumentar em número ou massa para serem distribuídos às células-filhas. Esse processo envolve replicação de DNA organelar, importação de proteínas, síntese de membranas, montagem de complexos e mecanismos de constrição e separação. Proteínas relacionadas a sistemas bacterianos de divisão foram adaptadas, modificadas ou substituídas ao longo da evolução, mas a lógica geral ainda preserva traços da fissão procariótica. A continuidade dessas organelas entre gerações celulares reforça a ideia de que a célula eucarionte é também uma linhagem de compartimentos herdados.
A dinâmica morfológica das mitocôndrias acrescenta outro nível de organização. Em muitas células, mitocôndrias não permanecem como estruturas isoladas e imutáveis; elas podem fundir-se, dividir-se, deslocar-se e formar redes. A fusão permite mistura de componentes, compensação de danos e manutenção funcional; a fissão facilita distribuição, renovação e remoção seletiva de partes danificadas por mitofagia. A forma da rede mitocondrial responde ao estado energético, ao ciclo celular, ao estresse e às necessidades de cada tipo celular. Em células vegetais, mitocôndrias geralmente são numerosas e móveis, deslocando-se em associação ao citoesqueleto e interagindo metabolicamente com cloroplastos e peroxissomos. Assim, a mitocôndria deve ser vista como uma população dinâmica de organelas, não como estruturas fixas desenhadas em cortes histológicos.
Os cloroplastos também apresentam dinâmica intracelular. Em células foliares, podem mudar de posição conforme a intensidade luminosa, otimizando a captura de luz sob condições limitantes ou reduzindo dano quando a radiação é excessiva. Esse reposicionamento depende do citoesqueleto, de sinais luminosos e de mecanismos celulares que ajustam a distribuição das organelas no interior da célula. Em uma folha exposta ao sol intenso do semiárido, por exemplo, a posição e o estado funcional dos cloroplastos influenciam a capacidade de equilibrar fotossíntese e fotoproteção. A organela fotossintética não é apenas uma estrutura bioquímica; é também um componente espacialmente regulado em uma célula submetida a variações ambientais rápidas. [Figura 6.19]
A relação entre mitocôndrias, cloroplastos e espécies reativas de oxigênio merece atenção especial. Como ambas as organelas realizam transporte de elétrons, ambas podem gerar espécies reativas em determinadas condições. Essas moléculas podem causar danos oxidativos a proteínas, lipídios e ácidos nucleicos, mas também atuam como sinais em concentrações controladas. A célula precisa equilibrar produção e remoção dessas espécies por meio de sistemas antioxidantes distribuídos entre organelas, citosol e outros compartimentos. Em plantas, situações como excesso de luz, fechamento estomático, seca, salinidade e deficiência nutricional podem alterar a pressão redox sobre cloroplastos e mitocôndrias. A tolerância ao estresse depende da capacidade de manter esse equilíbrio, coordenando metabolismo energético, defesa antioxidante e reprogramação gênica.
A semiautonomia dessas organelas, portanto, não significa isolamento. Mitocôndrias e cloroplastos dependem de proteínas nucleares, de metabólitos citosólicos, de lipídios produzidos ou distribuídos por outros compartimentos, de sistemas de qualidade proteica e de sinais regulatórios vindos de toda a célula. Ao mesmo tempo, influenciam o metabolismo global, a expressão gênica, o estado redox e a resposta ao ambiente. Essa relação bidirecional é central para a lógica eucarionte. O núcleo não governa sozinho de maneira unilateral; ele coordena parte importante da informação genética, mas recebe sinais constantes de organelas que informam sobre energia, metabolismo, estresse e desenvolvimento. A célula eucarionte é regulada por diálogo interno, não por comando linear simples.
A integração entre genoma nuclear e genomas organelares também oferece uma perspectiva importante sobre evolução. A endossimbiose não foi apenas uma associação temporária entre células diferentes. Ela envolveu transferência de genes, perda de autonomia, evolução de sistemas de importação proteica, ajuste de divisão organelar, compatibilização metabólica e desenvolvimento de mecanismos de comunicação. Esse processo transformou antigos organismos independentes em organelas indispensáveis. O resultado não é uma simples soma entre hospedeiro e simbionte, mas uma nova unidade biológica. A célula eucarionte moderna carrega, em sua própria organização, vestígios de cooperação evolutiva convertida em dependência estrutural e funcional.
Na perspectiva do ensino de biologia celular, mitocôndrias e cloroplastos são frequentemente tratados apenas nos capítulos de bioenergética, quando se estudam respiração celular e fotossíntese. Essa abordagem é necessária, mas insuficiente. Antes de discutir cadeias transportadoras de elétrons, ATP sintase, ciclo do ácido cítrico, fotossistemas ou ciclo de Calvin-Benson, é preciso reconhecer que essas organelas são componentes estruturais e evolutivos da organização eucarionte. Elas possuem membranas próprias, genomas próprios, divisão própria, importação proteica específica e comunicação com o núcleo. A bioenergética é apenas uma de suas dimensões, embora seja uma dimensão central. Sua presença reorganizou a escala de possibilidades da célula eucarionte.
Essa reorganização energética provavelmente contribuiu para a expansão da complexidade celular. Células maiores, genomas maiores, citoesqueleto dinâmico, tráfego vesicular, endocitose, secreção, diferenciação e multicelularidade complexa exigem energia e regulação. A disponibilidade de organelas especializadas em conversão energética permitiu que eucariontes explorassem formas de organização interna mais custosas e sofisticadas. Em plantas, a associação entre mitocôndrias e cloroplastos ampliou ainda mais essa capacidade, permitindo que a célula produzisse compostos orgânicos a partir de CO₂ e luz, ao mesmo tempo em que mantinha sistemas respiratórios para mobilizar energia conforme a demanda. A célula vegetal é, nesse sentido, um exemplo extremo de integração energética e compartimental.
A agricultura depende profundamente dessa integração. A produtividade de uma cultura não é apenas consequência da presença de cloroplastos, mas da eficiência com que eles capturam luz, fixam carbono, exportam açúcares, evitam dano oxidativo e comunicam seu estado ao restante da célula. A respiração mitocondrial, por sua vez, não representa simples consumo de fotoassimilados; ela fornece energia, intermediários metabólicos e flexibilidade fisiológica para crescimento, manutenção, absorção de nutrientes, resposta a estresses e desenvolvimento reprodutivo. Em sementes, raízes, frutos e tecidos jovens, mitocôndrias são decisivas mesmo quando a fotossíntese local é ausente ou limitada. Melhorar plantas para ambientes adversos exige considerar essa rede, porque tolerância, crescimento e rendimento resultam da coordenação entre captura de energia, uso de energia e proteção contra danos.
A biotecnologia vegetal explora direta ou indiretamente essa lógica. Estratégias para aumentar eficiência fotossintética, melhorar tolerância ao estresse oxidativo, alterar acúmulo de amido, modificar composição de óleos, produzir metabólitos de interesse ou expressar proteínas recombinantes podem envolver plastídios, mitocôndrias ou sua comunicação com o núcleo. A engenharia de cloroplastos, em particular, atrai interesse porque plastídios podem acumular altos níveis de determinadas proteínas e participar de vias metabólicas específicas. No entanto, qualquer intervenção nesse nível precisa considerar que a organela opera dentro de uma rede celular. Alterar um componente do cloroplasto pode afetar metabolismo, redox, desenvolvimento e sinalização. A compartimentalização oferece oportunidades, mas também impõe interdependências.
A presença de mitocôndrias e cloroplastos também mostra que a célula eucarionte não deve ser compreendida como uma estrutura inteiramente inventada de dentro para fora. Parte de sua complexidade resultou da incorporação, integração e transformação de outras células. Essa constatação muda a forma de pensar evolução celular. A inovação biológica não ocorre apenas por mutações graduais em uma linhagem isolada; pode envolver associação entre organismos, transferência de funções, reorganização de genomas e construção de novas unidades de seleção. A célula eucarionte é, em certo sentido, uma comunidade evolutiva estabilizada, na qual antigas individualidades foram convertidas em funções internas.
Ao reunir genoma nuclear, citosol, mitocôndrias e cloroplastos em uma mesma unidade funcional, a célula eucarionte criou uma arquitetura capaz de integrar informação, energia e metabolismo em múltiplos compartimentos. Mitocôndrias e cloroplastos conservaram parte de sua herança procariótica, mas foram profundamente remodelados pela vida dentro da célula hospedeira. Elas se dividem, expressam genes, produzem algumas proteínas e sustentam processos energéticos centrais, mas dependem do núcleo, do citosol e de sistemas de importação para existir. A semiautonomia dessas organelas é, portanto, uma autonomia regulada, construída ao longo da evolução e mantida por comunicação contínua. Na lógica eucarionte, mitocôndrias e cloroplastos não são apenas compartimentos de energia; são testemunhos vivos de uma história evolutiva em que cooperação, integração e dependência deram origem a novas possibilidades celulares.
6.6 — Peroxissomos e compartimentos oxidativos #
Entre as organelas da célula eucarionte, os peroxissomos costumam receber menos atenção do que mitocôndrias, cloroplastos, retículo endoplasmático ou núcleo. Essa diferença não decorre de menor importância biológica, mas de uma combinação de fatores históricos e didáticos. Os peroxissomos são menores, apresentam morfologia aparentemente simples, não possuem DNA próprio, não formam sistemas internos de membranas comparáveis às cristas mitocondriais ou aos tilacoides cloroplastidiais e, em muitos esquemas introdutórios, aparecem apenas como vesículas associadas à detoxificação. Essa representação é insuficiente. Os peroxissomos são compartimentos metabólicos altamente especializados, delimitados por membrana simples, ricos em enzimas oxidativas e integrados a processos essenciais como metabolismo lipídico, controle de espécies reativas de oxigênio, fotorrespiração, germinação de sementes, senescência, resposta a estresses e sinalização celular. [Figura 6.20]
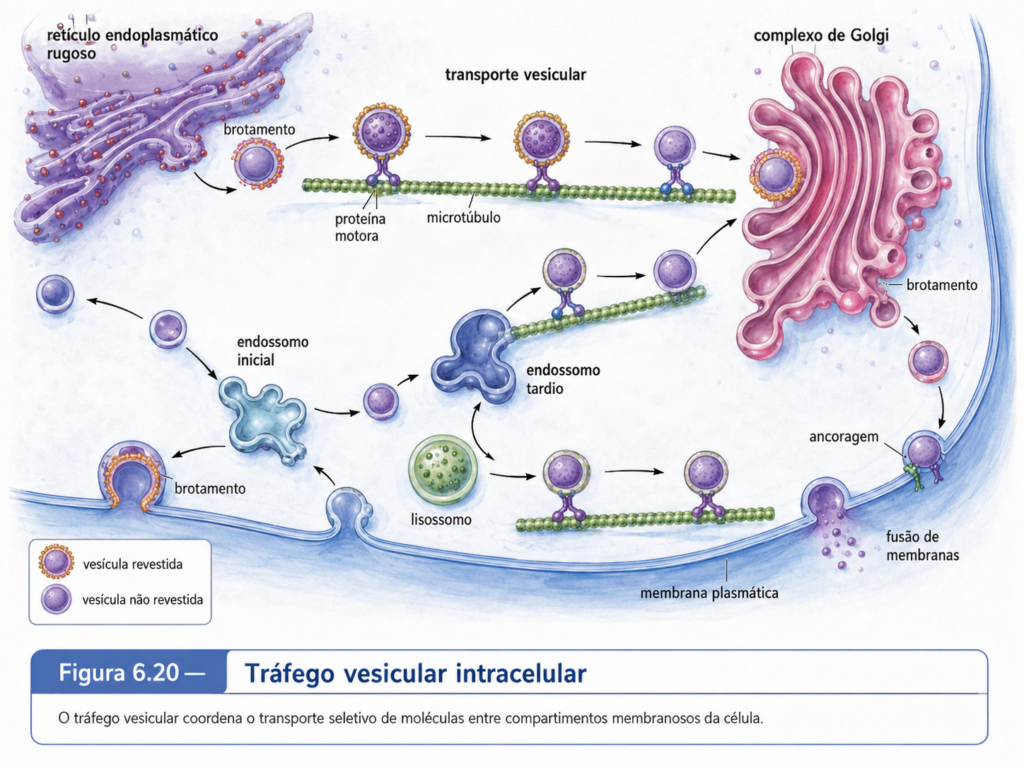
A própria denominação “peroxissomo” indica uma de suas características centrais: a relação com o peróxido de hidrogênio. Muitas enzimas presentes nessas organelas utilizam oxigênio molecular em reações de oxidação, transferindo elétrons de substratos orgânicos para o O₂ e formando H₂O₂ como produto. O peróxido de hidrogênio é uma molécula ambivalente. Em concentrações elevadas, pode contribuir para dano oxidativo, afetando proteínas, lipídios e ácidos nucleicos. Em concentrações controladas, pode atuar como sinal envolvido em respostas ao ambiente, desenvolvimento e defesa. A célula precisa, portanto, produzir, conter, utilizar e remover H₂O₂ de maneira regulada. Os peroxissomos resolvem parte desse problema reunindo enzimas produtoras de peróxido e enzimas capazes de degradá-lo no mesmo compartimento, especialmente a catalase, que converte H₂O₂ em água e oxigênio. [Figura 6.21]
A presença de catalase é uma das marcas funcionais mais conhecidas dos peroxissomos. Essa enzima permite que reações oxidativas ocorram sem que o peróxido de hidrogênio se acumule livremente no citosol em níveis incompatíveis com a homeostase celular. A compartimentalização tem aqui um significado claro: a célula não evita completamente reações perigosas; ela as coloca em locais apropriados, cercadas por sistemas de contenção. Em vez de dispersar oxidases pelo citosol, concentra parte delas em organelas especializadas e associa sua atividade a mecanismos antioxidantes. Essa organização permite explorar o potencial químico do oxigênio sem transformar o próprio metabolismo em ameaça permanente. [Tabela 6.6]
A membrana peroxissomal é essencial para essa lógica. Embora seja uma membrana simples, ela delimita um ambiente interno com composição enzimática própria e controla a entrada e saída de proteínas, metabólitos e produtos de reação. Diferentemente de mitocôndrias e cloroplastos, os peroxissomos não possuem genoma nem ribossomos. Todas as suas proteínas são codificadas no núcleo, sintetizadas em ribossomos citosólicos e direcionadas à organela por sinais específicos de endereçamento. Essa dependência do núcleo e do citosol mostra que o peroxissomo é integralmente eucariótico em sua biogênese atual, embora mantenha uma função metabólica altamente especializada. A organela não se sustenta por autonomia genética, mas por importação seletiva de proteínas e manutenção dinâmica de sua membrana.
O direcionamento de proteínas aos peroxissomos depende de sequências sinal reconhecidas por receptores citosólicos e por proteínas envolvidas na biogênese peroxissomal, frequentemente chamadas peroxinas. Uma característica notável desse sistema é a capacidade de importar algumas proteínas já dobradas ou mesmo associadas a cofatores, o que diferencia os peroxissomos de organelas que exigem a translocação de cadeias polipeptídicas mais desdobradas. Essa propriedade amplia a flexibilidade funcional da organela, permitindo que enzimas cataliticamente maduras sejam incorporadas ao seu interior. Ao mesmo tempo, exige controle rigoroso, pois a importação seletiva define a identidade metabólica do peroxissomo. Uma pequena organela pode mudar sua composição enzimática e, com isso, modificar seu papel fisiológico conforme o tipo celular, o estágio de desenvolvimento ou a condição ambiental. [Figura 6.22]
A biogênese dos peroxissomos não deve ser imaginada como um processo único e rígido. Eles podem crescer, dividir-se, receber lipídios e proteínas de diferentes fontes, estabelecer relações com o retículo endoplasmático e ajustar sua abundância conforme a demanda metabólica. Em determinadas condições, a célula aumenta o número de peroxissomos; em outras, reduz ou remove organelas excedentes por processos degradativos seletivos. Essa plasticidade é coerente com sua função. Como compartimentos ligados ao metabolismo oxidativo, os peroxissomos precisam responder rapidamente a mudanças em disponibilidade de substratos, estado redox, desenvolvimento e estresse. A célula não mantém uma população fixa de peroxissomos apenas por estabilidade estrutural; ela regula essas organelas conforme a necessidade funcional.
Em células animais, peroxissomos participam de processos como β-oxidação de ácidos graxos de cadeia muito longa, metabolismo de lipídios específicos, detoxificação de compostos e controle de espécies reativas de oxigênio. Algumas dessas funções são complementares às mitocondriais, mas não idênticas. A β-oxidação peroxissomal pode encurtar ácidos graxos muito longos, gerando produtos que depois serão metabolizados por outras vias. O metabolismo lipídico associado aos peroxissomos também se relaciona à formação de moléculas importantes para membranas e tecidos especializados. Alterações graves na biogênese peroxissomal ou em enzimas peroxissomais podem produzir doenças sistêmicas, evidenciando que essas organelas são pequenas apenas em tamanho, não em relevância fisiológica.
Nas células vegetais, a importância dos peroxissomos é ainda mais evidente quando se considera sua diversidade funcional. Eles participam da fotorrespiração, do metabolismo de ácidos graxos durante a germinação de sementes oleaginosas, da detoxificação de espécies reativas de oxigênio, da biossíntese e modulação de sinais hormonais, da senescência e de respostas a estresses ambientais. Em folhas fotossintéticas, os peroxissomos operam em estreita relação com cloroplastos e mitocôndrias. Durante a fotorrespiração, compostos produzidos no cloroplasto são metabolizados em etapas que envolvem também peroxissomos e mitocôndrias, formando uma rota multiorganelar. Essa integração mostra que o peroxissomo vegetal não é um compartimento isolado de detoxificação, mas um nó metabólico que conecta fotossíntese, respiração, metabolismo de carbono, nitrogênio e estado redox. [Figura 6.23]
A fotorrespiração ilustra com força a necessidade de abandonar a visão fragmentada das organelas. Quando a rubisco atua sobre O₂ em vez de CO₂, forma-se 2-fosfoglicolato, uma molécula que precisa ser recuperada por uma série de reações distribuídas entre cloroplasto, peroxissomo, mitocôndria e citosol. Esse processo consome energia e libera CO₂, sendo frequentemente apresentado como perda de eficiência fotossintética. No entanto, em condições naturais, especialmente sob alta temperatura, seca ou limitação de CO₂, a fotorrespiração também contribui para a proteção do aparato fotossintético e para o equilíbrio metabólico. Os peroxissomos participam dessa rota gerando e removendo H₂O₂, conectando metabolismo de carbono e controle oxidativo. Em plantas cultivadas sob condições de estresse, essa integração pode influenciar crescimento, produtividade e tolerância ambiental. [Quadro 6.6]
Durante a germinação de sementes oleaginosas, peroxissomos especializados, frequentemente chamados glioxissomos, assumem papel central na mobilização de reservas lipídicas. Os lipídios armazenados são degradados, e seus produtos podem alimentar vias que permitem a conversão de carbono lipídico em carboidratos, sustentando o crescimento inicial da plântula antes que a fotossíntese esteja plenamente estabelecida. Essa função é particularmente importante porque sementes em germinação precisam transformar reservas químicas em energia, precursores biossintéticos e estruturas celulares novas. O peroxissomo, nesse caso, participa diretamente da transição entre vida quiescente da semente e crescimento ativo da plântula. A organela não apenas elimina substâncias reativas; ela ajuda a reorganizar o metabolismo em momentos críticos do desenvolvimento vegetal.
A plasticidade dos peroxissomos vegetais permite que sua função varie conforme o tecido. Em folhas maduras, podem estar fortemente associados à fotorrespiração. Em sementes germinantes, ao metabolismo de lipídios. Em tecidos submetidos a estresse, ao controle oxidativo e à sinalização. Em células senescentes, à remobilização metabólica e ao equilíbrio entre degradação e sobrevivência. Essa diversidade funcional reflete mudanças no conjunto de enzimas importadas para a organela. Como todas essas proteínas são codificadas no núcleo, a célula pode alterar a identidade bioquímica dos peroxissomos por mudanças na expressão gênica, tradução, importação proteica e proliferação organelar. O peroxissomo é, portanto, uma plataforma ajustável de metabolismo oxidativo.
A relação entre peroxissomos e espécies reativas de oxigênio exige uma interpretação equilibrada. Durante muito tempo, moléculas como H₂O₂ foram tratadas quase exclusivamente como subprodutos tóxicos. Essa visão foi substituída por uma compreensão mais refinada: espécies reativas podem causar dano, mas também funcionam como sinais. O que define seu efeito é a concentração, o local de produção, a duração do sinal, a presença de antioxidantes e a sensibilidade das moléculas-alvo. Os peroxissomos contribuem para essa regulação porque produzem e degradam H₂O₂ em um compartimento específico, permitindo sinalização localizada ou controlada. Em plantas, o H₂O₂ peroxissomal pode participar de respostas a patógenos, fechamento estomático, senescência, estresse salino, estresse hídrico e alta luminosidade, sempre em interação com sinais vindos de cloroplastos, mitocôndrias, membrana plasmática e núcleo.
Essa interação com outras organelas é uma das dimensões mais importantes da biologia peroxissomal. Peroxissomos mantêm proximidade funcional com mitocôndrias em processos de metabolismo lipídico, equilíbrio redox e degradação de ácidos graxos. Em células vegetais, também se associam metabolicamente a cloroplastos durante a fotorrespiração e respostas à luz. O retículo endoplasmático contribui para aspectos da biogênese e manutenção peroxissomal. O citosol fornece proteínas, metabólitos, cofatores e sistemas regulatórios. O núcleo ajusta a expressão dos genes necessários ao funcionamento da organela. A identidade do peroxissomo resulta dessa rede. Ele é delimitado por membrana, mas funcionalmente aberto à célula inteira.
A proximidade física entre organelas pode favorecer essa integração. Em muitas células, peroxissomos se deslocam pelo citoplasma associados ao citoesqueleto e podem aproximar-se de mitocôndrias, cloroplastos, gotas lipídicas e retículo endoplasmático. Essa dinâmica facilita trocas metabólicas e respostas rápidas. Em sementes oleaginosas, por exemplo, a proximidade entre peroxissomos e corpos lipídicos favorece a mobilização de reservas. Em células fotossintéticas, a relação espacial entre cloroplastos, peroxissomos e mitocôndrias contribui para a eficiência da fotorrespiração. A célula organiza sua bioquímica também por posicionamento espacial, não apenas por presença ou ausência de enzimas.
Os peroxissomos também participam do metabolismo de compostos relacionados a hormônios e sinais vegetais. Algumas etapas da biossíntese de jasmonatos, moléculas envolvidas em defesa, desenvolvimento e resposta a ferimentos, ocorrem em associação com peroxissomos. Essa conexão amplia o papel da organela para além do metabolismo energético ou antioxidante. Quando uma planta sofre ataque de herbívoros ou patógenos, sinais lipídicos e rotas oxidativas podem contribuir para respostas de defesa sistêmica e local. O peroxissomo participa dessa conversão entre dano percebido, metabolismo lipídico e sinalização hormonal. A organela, nesse caso, atua como interface entre metabolismo e decisão fisiológica. [Figura 6.24]
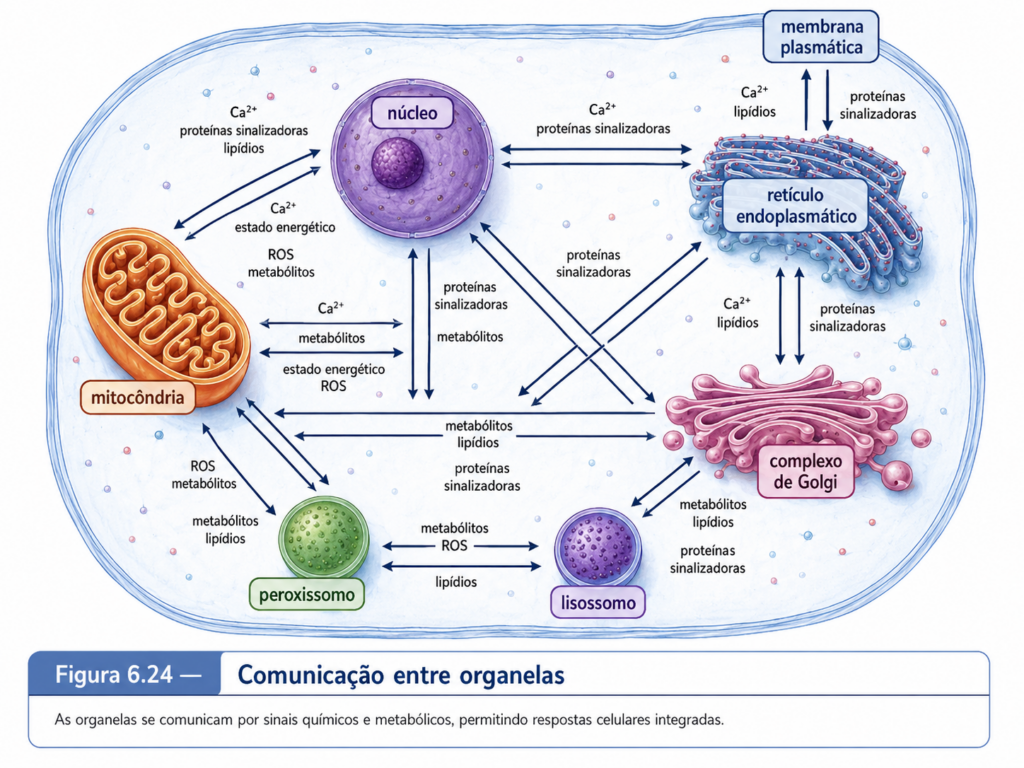
Na agricultura, essa dimensão é relevante porque tolerância a estresses raramente depende de uma única via. Plantas expostas à seca frequentemente enfrentam, ao mesmo tempo, fechamento estomático, redução de CO₂ interno, excesso relativo de energia luminosa nos cloroplastos, aumento de espécies reativas, mudanças hormonais e reorganização do metabolismo. A salinidade adiciona toxicidade iônica e estresse osmótico. O calor afeta membranas, proteínas e eficiência enzimática. Em todos esses cenários, peroxissomos participam do equilíbrio oxidativo e metabólico, funcionando em rede com cloroplastos, mitocôndrias, vacúolos e citosol. Melhoramento genético, manejo nutricional e uso de bioestimulantes que buscam aumentar tolerância ao estresse acabam interferindo, direta ou indiretamente, em sistemas celulares nos quais os peroxissomos estão envolvidos.
A compreensão dos peroxissomos também ajuda a interpretar a ação de antioxidantes e sistemas de defesa celular. A célula não possui um único mecanismo antioxidante universal. Ela distribui enzimas e moléculas redutoras entre compartimentos: catalase nos peroxissomos, ascorbato peroxidases em diferentes regiões celulares, superóxido dismutases em organelas e citosol, glutationa e ascorbato em redes redox integradas. Essa distribuição é importante porque espécies reativas são produzidas em locais específicos. Não basta ter antioxidantes em qualquer lugar; é necessário que o sistema de controle esteja próximo ou funcionalmente conectado ao local de geração do sinal ou do dano. O peroxissomo representa um desses centros de controle localizado.
A investigação dos peroxissomos avançou por meio de uma combinação de microscopia, citoquímica, fracionamento celular, genética e biologia molecular. A localização da catalase e de oxidases em pequenas organelas delimitadas por membrana foi decisiva para reconhecer esses compartimentos como entidades funcionais. A microscopia eletrônica permitiu observar sua morfologia; reações citoquímicas evidenciaram atividade enzimática; o fracionamento celular permitiu associar frações subcelulares a enzimas oxidativas; mutantes com defeitos em biogênese peroxissomal revelaram proteínas necessárias à formação e importação de componentes. Essa trajetória experimental mostra novamente que a biologia celular amadurece quando estrutura e função são analisadas juntas. Uma organela não é definida apenas por ser visível, mas por concentrar processos específicos demonstráveis.
Em células humanas, a relevância dos peroxissomos é evidenciada por doenças associadas à falha em sua biogênese ou em enzimas específicas. Quando proteínas necessárias à importação peroxissomal são defeituosas, múltiplas enzimas deixam de alcançar a organela, comprometendo várias vias ao mesmo tempo. O resultado pode ser grave, afetando sistema nervoso, fígado, rins, desenvolvimento e metabolismo lipídico. Essas condições mostram que uma organela aparentemente pequena pode sustentar funções sistêmicas indispensáveis. Embora o foco do livro dialogue fortemente com biologia vegetal e ciências agrárias, a perspectiva comparativa ajuda a reforçar um princípio geral: compartimentos celulares especializados mantêm processos cuja falha repercute no organismo inteiro.
Em plantas, defeitos peroxissomais também podem comprometer germinação, desenvolvimento, fotossíntese, resposta hormonal e resistência ao estresse. Mutantes com alterações em enzimas da fotorrespiração, da β-oxidação ou da biogênese peroxissomal frequentemente apresentam crescimento reduzido, sensibilidade a condições ambientais ou problemas em fases específicas do desenvolvimento. Isso reforça que os peroxissomos não são acessórios metabólicos. Eles participam de rotas que conectam reservas, energia, carbono, nitrogênio, lipídios, sinais hormonais e controle redox. A célula vegetal depende dessa organela para ajustar sua fisiologia em ambientes variáveis.
A ausência de DNA nos peroxissomos diferencia essas organelas de mitocôndrias e cloroplastos, mas também evidencia outro modelo de especialização eucariótica. Mitocôndrias e cloroplastos carregam uma história endossimbiótica marcada por genomas próprios. Peroxissomos, por outro lado, mostram como a célula pode construir um compartimento funcional inteiramente dependente do sistema nuclear-citosólico de síntese e importação proteica. Sua identidade não vem de um genoma interno, mas de sinais de endereçamento, receptores, membrana seletiva, composição enzimática e integração metabólica. A célula eucarionte, portanto, combina diferentes estratégias de construção de organelas: algumas derivadas de endossimbiose, outras formadas por redes internas de membranas e importação proteica.
A multiplicação e a remoção seletiva de peroxissomos acrescentam outra camada de regulação. Quando há necessidade de maior capacidade oxidativa ou metabolismo lipídico, a célula pode aumentar a abundância dessas organelas. Quando peroxissomos se tornam danificados ou excedentes, podem ser degradados por pexofagia, uma forma seletiva de autofagia. Esse controle quantitativo impede tanto a deficiência quanto o excesso de compartimentos oxidativos. Como peroxissomos produzem H₂O₂, sua proliferação descontrolada ou sua remoção ineficiente poderia alterar o equilíbrio redox celular. A célula precisa ajustar não apenas a atividade enzimática interna, mas também o número, a distribuição e a qualidade dessas organelas.
O peroxissomo revela, assim, um princípio importante da organização eucarionte: a célula não compartimentaliza apenas processos grandes ou visualmente complexos. Ela compartimentaliza também reações químicas específicas cujo controle espacial oferece vantagem funcional. Reações oxidativas poderiam ocorrer no citosol, mas sua concentração em uma organela permite maior eficiência, menor dano colateral e integração com sistemas antioxidantes. Metabolismo de lipídios, fotorrespiração, sinalização por H₂O₂ e resposta a estresse tornam-se mais controláveis quando organizados em um ambiente próprio. A organela funciona como uma zona de metabolismo intensivo, delimitada o suficiente para conter riscos e conectada o suficiente para integrar-se à célula.
Essa visão evita dois extremos. O primeiro seria tratar os peroxissomos como simples “depósitos de catalase”, reduzindo sua diversidade funcional. O segundo seria isolá-los artificialmente das demais organelas, como se suas funções pudessem ser compreendidas sem mitocôndrias, cloroplastos, retículo endoplasmático, citosol e núcleo. A interpretação correta está na integração. Peroxissomos concentram reações oxidativas e metabólicas específicas, mas seus substratos, produtos, proteínas e sinais vêm de outras partes da célula e retornam a elas. São compartimentos especializados em uma rede maior.
A importância dos peroxissomos torna-se ainda mais clara quando se pensa em ambientes agrícolas adversos. Em regiões semiáridas, plantas cultivadas frequentemente enfrentam ciclos de seca, calor, alta irradiância, salinidade localizada e limitações nutricionais. Essas condições afetam diretamente cloroplastos e mitocôndrias, mas também exigem forte participação dos peroxissomos na contenção oxidativa e no ajuste metabólico. Uma planta que mantém equilíbrio redox, recicla metabólitos, mobiliza reservas e coordena sinais hormonais tem maior chance de preservar crescimento e produção. A organela pequena, quase discreta nos esquemas, participa silenciosamente de processos que definem desempenho agronômico.
A célula eucarionte, portanto, utiliza peroxissomos como compartimentos oxidativos flexíveis, capazes de concentrar enzimas, processar metabólitos, controlar peróxido de hidrogênio, interagir com outras organelas e ajustar respostas fisiológicas. Sua simplicidade morfológica aparente contrasta com sua sofisticação funcional. Eles mostram que a complexidade celular não depende apenas de estruturas grandes ou visualmente elaboradas, mas da capacidade de organizar reações no espaço, controlar riscos químicos e integrar fluxos metabólicos. Na arquitetura eucariótica, os peroxissomos ocupam o lugar dos compartimentos que transformam oxidação em função, perigo químico em sinal regulado e metabolismo localizado em resposta celular integrada.
6.7 — Citoesqueleto e arquitetura dinâmica da célula eucarionte #
A compartimentalização da célula eucarionte criou um problema organizacional que não existia na mesma escala nas células menores e mais simples: como manter ordem interna em um volume celular maior, repleto de organelas, vesículas, ribossomos, complexos proteicos, regiões especializadas de membrana e fluxos constantes de moléculas. A presença de compartimentos não bastaria se eles permanecessem distribuídos ao acaso. Mitocôndrias precisam ocupar regiões compatíveis com a demanda energética; vesículas precisam alcançar membranas específicas; cromossomos precisam ser segregados durante a divisão celular; a forma celular precisa ser mantida ou modificada; células precisam migrar, crescer, alongar-se, dividir-se ou estabelecer polaridade. O citoesqueleto responde a esse desafio. Ele transforma o citoplasma em um espaço organizado, mecânico e dinâmico, no qual estruturas internas podem ser posicionadas, deslocadas, sustentadas e remodeladas conforme as necessidades da célula. [Figura 6.25]
O termo citoesqueleto pode sugerir uma armação rígida, comparável ao esqueleto ósseo de um animal. Essa imagem é inadequada. O citoesqueleto celular é uma rede de polímeros proteicos altamente dinâmica, capaz de montar-se, desmontar-se, reorganizar-se e interagir com membranas, organelas, proteínas motoras e sinais bioquímicos. Ele não apenas sustenta a célula; participa do movimento, da divisão, do tráfego intracelular, da organização espacial, da resposta mecânica e da definição da polaridade celular. Suas principais classes estruturais são os microfilamentos de actina, os microtúbulos e os filamentos intermediários. Cada uma dessas redes possui propriedades próprias de composição, diâmetro, rigidez, polaridade, estabilidade e interação com proteínas associadas, mas suas funções se sobrepõem e se integram em muitos processos celulares. [Tabela 6.7]
Os microfilamentos de actina formam estruturas finas, flexíveis e altamente remodeláveis, frequentemente associadas à periferia celular, à contração, à movimentação da membrana plasmática, à formação de projeções celulares, ao tráfego de curta distância e à divisão celular. Em células animais, a actina participa de lamelipódios, filopódios, microvilosidades, anéis contráteis e regiões de adesão. Em células vegetais, filamentos de actina estão fortemente relacionados ao movimento citoplasmático, ao posicionamento de organelas, ao crescimento polarizado e à organização de processos de secreção. Essa rede é sensível a sinais celulares e pode ser rapidamente reorganizada em resposta a estímulos externos, estado fisiológico, mudanças hormonais, ferimentos ou interações com patógenos. A actina confere à célula capacidade de reorganização local, permitindo que determinadas regiões da membrana cresçam, retraiam-se, secretem componentes ou respondam ao ambiente.
Os microtúbulos, por sua vez, são polímeros mais rígidos, formados por tubulina, que atuam como elementos estruturais e trilhos para transporte intracelular. Eles contribuem para o posicionamento de organelas, a organização do citoplasma, a formação do fuso mitótico e meiótico, o transporte de vesículas e, em muitas células, a formação de cílios e flagelos. Sua polaridade estrutural permite que proteínas motoras reconheçam direções distintas e transportem cargas para regiões específicas. Durante a divisão celular, os microtúbulos reorganizam-se profundamente para capturar, alinhar e separar cromossomos. Em células vegetais, embora não haja centríolos típicos na maioria das células superiores, microtúbulos corticais orientam a deposição de celulose na parede celular, influenciando a direção de expansão celular e, consequentemente, a forma dos tecidos. [Figura 6.26]
Os filamentos intermediários formam uma rede mais resistente e estável, especialmente importante em células animais submetidas a tensão mecânica. Eles contribuem para a integridade estrutural de células epiteliais, neurônios, fibras musculares e do núcleo, onde as lâminas nucleares sustentam o envoltório nuclear e participam da organização da cromatina. Embora plantas não possuam filamentos intermediários citoplasmáticos clássicos equivalentes aos de animais, apresentam proteínas estruturais e sistemas de sustentação próprios que, junto à parede celular, ao citoesqueleto de actina e aos microtúbulos, contribuem para resistência e forma. A comparação entre células animais e vegetais mostra que a sustentação celular pode ser resolvida por estratégias distintas: em animais, a matriz extracelular, adesões e redes internas de filamentos exercem papel central; em plantas, a parede celular acrescenta uma dimensão mecânica externa indispensável.
A importância do citoesqueleto torna-se evidente quando se abandona a ideia de citoplasma como meio homogêneo. Em uma célula viva, organelas não ficam simplesmente flutuando em uma solução aquosa. Elas podem ser ancoradas, transportadas, reposicionadas ou distribuídas por interações com filamentos e proteínas motoras. Mitocôndrias podem deslocar-se para regiões de maior demanda energética; vesículas secretoras podem ser direcionadas a áreas específicas da membrana plasmática; endossomos podem trafegar entre periferia e regiões internas; cloroplastos podem mudar de posição em resposta à intensidade luminosa; vacúolos e peroxissomos podem assumir distribuições compatíveis com o estado metabólico da célula. A arquitetura celular não é uma fotografia fixa. É uma organização em movimento, sustentada por redes proteicas que respondem ao tempo, ao espaço e ao sinal. [Animação 6.4]
O transporte intracelular dirigido é uma das funções mais importantes do citoesqueleto. Em células pequenas, a difusão pode ser suficiente para muitos processos; em células eucariontes maiores, polarizadas ou altamente especializadas, depender apenas da difusão seria lento e impreciso. Proteínas motoras resolvem esse problema convertendo energia química, geralmente da hidrólise de ATP, em movimento mecânico. Cinesinas e dineínas deslocam cargas ao longo de microtúbulos, frequentemente em direções opostas; miosinas deslocam cargas ao longo de filamentos de actina. Essas cargas podem incluir vesículas, organelas, RNAs, complexos proteicos e componentes de membrana. A célula, portanto, possui uma logística interna baseada em trilhos, motores e endereços moleculares. Essa logística permite que a compartimentalização funcione de modo integrado, evitando que a distância interna comprometa a eficiência celular.
Em neurônios, essa exigência é extrema. Um axônio pode alcançar grande comprimento, e componentes sintetizados no corpo celular precisam ser transportados a regiões distantes, enquanto sinais e materiais também retornam em direção ao núcleo. Sem microtúbulos, motores moleculares e sistemas de tráfego associados, a polaridade neuronal seria inviável. Em células epiteliais, proteínas precisam ser direcionadas a domínios apicais ou basolaterais da membrana. Em células secretoras, vesículas precisam seguir rotas específicas até regiões de liberação. Em células vegetais, vesículas contendo componentes da parede celular devem ser entregues a locais de crescimento, divisão ou remodelamento. Esses exemplos mostram que o citoesqueleto não atua apenas em células “móveis”; atua em qualquer célula que precise distribuir componentes com precisão.
A relação entre citoesqueleto e membranas é particularmente estreita. A membrana plasmática não mantém sua forma e sua organização apenas por propriedades da bicamada lipídica. Ela interage com redes corticais de actina, proteínas adaptadoras, complexos de adesão, receptores e componentes da matriz extracelular ou da parede celular. Essa interação permite que sinais externos modifiquem a organização interna da célula e que forças internas alterem a superfície celular. Quando uma célula animal migra, a actina promove protrusão em uma extremidade, adesões se formam e se desfazem, miosinas geram contração e a membrana é continuamente remodelada por exocitose e endocitose. Quando uma célula vegetal cresce, microtúbulos corticais orientam a deposição de microfibrilas de celulose, enquanto actina e tráfego vesicular ajudam a entregar material de membrana e parede. Em ambos os casos, forma celular resulta de integração entre membrana, citoesqueleto e matriz externa.
A polaridade celular depende dessa integração. Uma célula polarizada possui regiões estrutural e funcionalmente distintas. Essa polaridade pode ser temporária, como em uma célula migrando em direção a um sinal, ou estável, como em células epiteliais, neurônios, tubos polínicos, pelos radiculares e células em diferenciação. Para estabelecer polaridade, a célula precisa distribuir proteínas, lipídios, organelas, RNAs e vesículas de maneira assimétrica. O citoesqueleto fornece trilhos e regiões de ancoragem para essa distribuição, enquanto sinais intracelulares definem onde a montagem de filamentos deve ser favorecida ou reprimida. Em plantas, o crescimento polarizado do tubo polínico é um exemplo notável: a expansão ocorre em uma extremidade, sustentada por intensa secreção vesicular, organização de actina, gradientes iônicos e controle da parede celular. [Figura 6.27]
O citoesqueleto também participa da divisão celular, um dos momentos em que a organização interna precisa ser mais radicalmente remodelada. Durante a mitose, microtúbulos formam o fuso mitótico, estrutura responsável por capturar cromossomos, alinhar cromátides e separá-las em direção aos polos celulares. Em células animais, a citocinese envolve um anel contrátil de actina e miosina que estrangula a célula em divisão. Em células vegetais, a separação das células-filhas depende da formação da placa celular, orientada por estruturas derivadas do citoesqueleto e pelo tráfego vesicular dirigido ao centro da célula. A divisão celular, portanto, não é apenas duplicação de DNA seguida de separação física. É um processo de engenharia intracelular no qual cromossomos, microtúbulos, actina, membranas, vesículas e parede celular precisam atuar em sequência coordenada.
A organização do citoesqueleto é regulada por proteínas associadas que controlam nucleação, alongamento, encurtamento, ramificação, estabilização, corte, ancoragem e interação com membranas. Filamentos de actina e microtúbulos não se organizam espontaneamente de maneira útil apenas por suas propriedades químicas básicas. A célula utiliza complexos nucleadores, proteínas de capeamento, proteínas de ligação lateral, proteínas estabilizadoras e fatores que promovem desmontagem. Esse controle permite que a rede seja adaptável. Uma região celular pode promover polimerização de actina enquanto outra desmonta filamentos; microtúbulos podem crescer explorando o citoplasma e desaparecer quando não são estabilizados; estruturas transitórias podem surgir durante a divisão e desaparecer depois. A célula economiza material e ganha plasticidade ao montar estruturas no momento e no local necessários.
A instabilidade dinâmica dos microtúbulos é um exemplo elegante dessa lógica. Microtúbulos podem alternar fases de crescimento e encurtamento, explorando o espaço celular até serem estabilizados por interações adequadas. Durante a mitose, essa propriedade permite que microtúbulos procurem cinetócoros nos cromossomos e estabeleçam conexões funcionais. Durante a organização do citoplasma, permite reposicionamento rápido conforme sinais celulares. A actina também apresenta dinâmica própria, com polimerização preferencial em uma extremidade, despolimerização em outra e formação de estruturas ramificadas ou feixes paralelos conforme as proteínas associadas. Esses comportamentos mostram que o citoesqueleto é um sistema fora do equilíbrio, sustentado por energia, regulação e renovação contínua.
A descoberta das proteínas motoras e da dinâmica dos filamentos transformou a compreensão da célula eucarionte. Durante muito tempo, movimentos intracelulares eram observados ao microscópio, mas seus mecanismos permaneciam obscuros. A combinação de microscopia, bioquímica, purificação de proteínas, ensaios in vitro e marcação fluorescente permitiu demonstrar que moléculas específicas podiam caminhar sobre filamentos, deslocando cargas. Experimentos com actina e miosina revelaram princípios de contração e movimento; estudos com microtúbulos permitiram identificar motores como cinesina e dineína; análises de células vivas mostraram organelas e vesículas trafegando por rotas definidas. A célula passou a ser vista como uma entidade mecanicamente ativa, não apenas como um saco químico compartimentalizado. [Figura 6.28]
Essa dimensão mecânica tem implicações profundas. Células percebem forças, respondem a rigidez do ambiente, modificam adesões, alteram expressão gênica e reorganizam seu citoesqueleto conforme estímulos físicos. Em tecidos animais, a tensão mecânica influencia forma, migração, diferenciação e organização tecidual. Em plantas, a parede celular impõe restrições e possibilidades específicas, e o citoesqueleto participa da orientação do crescimento em relação a tensões, luz, gravidade, água e contato. A célula não responde apenas a moléculas sinalizadoras; responde também a forças. O citoesqueleto é uma das principais vias pelas quais sinais mecânicos são convertidos em organização estrutural e respostas bioquímicas.
Nas células vegetais, a relação entre microtúbulos corticais e parede celular é decisiva para a morfogênese. A orientação dos microtúbulos influencia a deposição das microfibrilas de celulose, e a orientação da celulose condiciona a direção em que a célula pode expandir-se. Quando a expansão é mais restrita em uma direção e mais permissiva em outra, a célula alonga-se de modo orientado. Esse princípio, repetido em muitas células, contribui para a formação de tecidos, órgãos e arquitetura vegetal. O caule que se alonga, a raiz que penetra o solo, o pelo radicular que explora uma região específica e o tubo polínico que cresce em direção ao óvulo dependem de mecanismos celulares nos quais citoesqueleto, parede, membrana e secreção atuam de maneira integrada. [Quadro 6.7]
O movimento citoplasmático em células vegetais reforça a importância da actina. Em muitas células grandes, correntes citoplasmáticas transportam organelas e distribuem metabólitos ao longo de trajetórias internas. Esse fenômeno, visível em algumas células vegetais com relativa facilidade ao microscópio, depende de interações entre filamentos de actina, miosinas e cargas citoplasmáticas. Ele ajuda a superar limitações impostas pelo tamanho celular e pela presença de um grande vacúolo central. Quando o citosol fica comprimido na periferia ou em cordões citoplasmáticos, o movimento dirigido contribui para manter comunicação funcional entre regiões celulares. O citoesqueleto, nesse contexto, é parte da solução física para a vida em células grandes e vacuoladas.
A distribuição de cloroplastos também pode depender do citoesqueleto. Sob baixa intensidade luminosa, cloroplastos tendem a posicionar-se de modo a maximizar a captação de luz; sob luz excessiva, podem redistribuir-se para reduzir danos fotooxidativos. Esse movimento não é apenas uma curiosidade celular. Em ambientes agrícolas com alta irradiância, calor e déficit hídrico, a capacidade de ajustar a exposição dos cloroplastos pode contribuir para fotoproteção. O reposicionamento organelar, a dissipação de excesso de energia, o controle antioxidante e a comunicação entre cloroplastos e núcleo fazem parte de uma resposta integrada. A arquitetura interna da célula participa diretamente da adaptação da planta ao ambiente.
O citoesqueleto também está envolvido em interações entre plantas e patógenos. Durante a infecção, células vegetais podem reorganizar actina e microtúbulos para direcionar secreção de compostos de defesa, reforçar regiões da parede celular ou modular a entrada de invasores. Alguns patógenos, por sua vez, manipulam o citoesqueleto do hospedeiro para facilitar colonização. Essa disputa mostra que o citoesqueleto não é apenas uma infraestrutura interna neutra; é parte ativa da imunidade, da defesa e da interação ecológica. Em termos agronômicos, resistência a doenças envolve não apenas genes de reconhecimento e compostos antimicrobianos, mas também a capacidade celular de reorganizar rapidamente sua arquitetura e seu tráfego interno.
Em células animais, o citoesqueleto participa de processos igualmente críticos, como migração celular, cicatrização, resposta imune, contração muscular, formação de sinapses e manutenção da integridade epitelial. Leucócitos dependem de remodelamento de actina para migrar em direção a sinais inflamatórios. Células musculares utilizam arranjos altamente organizados de actina e miosina para gerar contração. Neurônios dependem de microtúbulos para transporte axonal e de actina para dinâmica de sinapses e cones de crescimento. Células epiteliais dependem de filamentos intermediários e adesões para resistir a tensões. Essas funções serão aprofundadas posteriormente, mas já evidenciam que a arquitetura dinâmica da célula eucarionte sustenta funções em escala de tecidos e organismos.
A relação entre citoesqueleto e núcleo é outro aspecto importante. O núcleo não está simplesmente solto no citoplasma. Sua posição pode ser controlada por microtúbulos, actina, motores moleculares e conexões com a lâmina nuclear. Em células em migração, divisão ou diferenciação, o posicionamento nuclear influencia polaridade e organização interna. Durante a divisão celular, microtúbulos interagem indiretamente com cromossomos por meio dos cinetócoros; durante a interfase, conexões mecânicas podem transmitir forças do citoplasma ao núcleo. Essas relações sugerem que a expressão gênica e a organização nuclear podem ser influenciadas por estados mecânicos e estruturais da célula. A separação entre arquitetura e regulação genética, portanto, é menos rígida do que muitos esquemas indicam.
O citoesqueleto também contribui para a organização de domínios celulares. Em células epiteliais, a separação entre regiões apical e basolateral depende de junções, tráfego vesicular e redes corticais. Em neurônios, axônio e dendritos possuem composições e funções distintas, sustentadas por organização diferencial de microtúbulos, actina e proteínas associadas. Em células vegetais, domínios de crescimento polarizado dependem de concentração localizada de vesículas, actina, cálcio, prótons e componentes de parede. A célula eucarionte pode, assim, criar regiões funcionais dentro de si mesma, sem precisar delimitá-las sempre por membranas internas completas. O citoesqueleto participa dessa compartimentalização espacial sem membrana.
Essa capacidade de criar assimetria é essencial para o desenvolvimento multicelular. Tecidos não se formam apenas porque células se dividem; formam-se porque células dividem-se em posições adequadas, crescem em direções específicas, aderem seletivamente, secretam matriz ou parede, respondem a gradientes e diferenciam domínios. O citoesqueleto participa de todos esses níveis. Em embriões animais, divisões assimétricas podem distribuir componentes celulares de modo desigual entre células-filhas. Em plantas, planos de divisão e direção de expansão ajudam a definir padrões de tecidos. A forma do organismo começa, em parte, com decisões físicas e espaciais tomadas dentro de células individuais.
A integração entre citoesqueleto e sistema de endomembranas é particularmente importante. Vesículas não apenas brotam e se fundem; precisam chegar ao local correto. O tráfego do retículo ao Golgi, do Golgi à membrana plasmática, da membrana aos endossomos e dos endossomos a compartimentos degradativos depende, em muitos casos, de transporte ao longo de filamentos. Durante secreção polarizada, crescimento celular ou resposta imune, essa coordenação se intensifica. Em células vegetais em crescimento, vesículas contendo componentes de parede precisam ser entregues a regiões específicas, e essa entrega depende de actina, microtúbulos e sinais locais. O sistema de endomembranas fornece membranas e cargas; o citoesqueleto fornece direção, força e organização espacial.
A dinâmica do citoesqueleto também depende do estado metabólico da célula. Polimerização, despolimerização, atividade de motores e manutenção de gradientes exigem energia. ATP e GTP participam diretamente da dinâmica de actina, microtúbulos e motores moleculares. Isso conecta citoesqueleto a mitocôndrias, cloroplastos e metabolismo citosólico. Uma célula com déficit energético pode alterar transporte, forma, divisão e secreção. Em plantas submetidas a estresse, mudanças na disponibilidade de energia e no estado redox podem afetar reorganização do citoesqueleto, crescimento de raízes, abertura estomática, expansão celular e respostas de defesa. A arquitetura celular é, portanto, dependente de energia, e a energia precisa ser distribuída em uma arquitetura funcional.
A ação de substâncias que interferem no citoesqueleto também ajudou a revelar sua importância. Compostos capazes de desestabilizar microtúbulos ou filamentos de actina alteram divisão celular, transporte intracelular, forma e movimento. Na medicina, fármacos que afetam microtúbulos são usados em contextos como tratamento de câncer, justamente porque células em divisão dependem do fuso mitótico. Na agricultura, herbicidas que interferem na organização de microtúbulos podem comprometer divisão e crescimento vegetal. Esses exemplos mostram que o citoesqueleto é um alvo funcional sensível: perturbar sua dinâmica pode bloquear processos celulares fundamentais. A utilidade aplicada desses compostos nasce da dependência profunda da célula eucarionte em relação à sua arquitetura interna. [Quadro 6.8]
A organização do citoesqueleto precisa ser estudada com métodos que capturem movimento. Imagens fixas são úteis para revelar arranjos estruturais, mas não mostram completamente a velocidade com que filamentos crescem, desaparecem, transportam cargas ou reorganizam a célula. A marcação de proteínas do citoesqueleto com sondas fluorescentes e proteínas fluorescentes permitiu observar filamentos em células vivas. Técnicas de microscopia de fluorescência, confocal, super-resolução e vídeo-microscopia revelaram a célula como um sistema em atividade contínua. O citoesqueleto tornou-se um dos exemplos mais claros de que a célula eucarionte deve ser compreendida no tempo, não apenas no espaço. [Vídeo 6.1]
O caráter dinâmico do citoesqueleto não significa ausência de controle. Pelo contrário, sua flexibilidade exige regulação rigorosa. Se filamentos se montassem em qualquer lugar, a célula perderia organização. Se fossem estáveis demais, a célula perderia adaptação. Se motores transportassem cargas indiscriminadamente, destinos celulares seriam confundidos. A vida celular depende de um equilíbrio entre estabilidade e mudança. O citoesqueleto precisa sustentar a forma, mas permitir remodelamento; precisa transportar cargas, mas preservar direção; precisa resistir a forças, mas transmitir sinais; precisa organizar divisão, mas desaparecer ou reorganizar-se ao final dela. Essa combinação de robustez e plasticidade é uma das marcas da célula eucarionte.
Em células vegetais, essa plasticidade precisa funcionar dentro de uma restrição adicional: a parede celular. Diferentemente de muitas células animais, que podem mudar de forma por deformação direta da membrana e do córtex celular, a célula vegetal cresce pressionando e remodelando uma parede externa. O turgor fornece força, a parede define resistência, e o citoesqueleto participa da orientação do crescimento e da entrega de novos materiais. Essa relação faz com que a forma vegetal resulte de uma negociação entre pressão interna, composição da parede, orientação de celulose, secreção vesicular e organização citosquelética. O alongamento de uma raiz, a expansão de uma folha ou a formação de vasos condutores dependem de processos celulares mecânicos e direcionais, não apenas de divisão e aumento de massa.
O citoesqueleto também ajuda a explicar por que a célula eucarionte pode assumir tantas formas. Um eritrócito, um neurônio, uma célula epitelial, uma fibra muscular, uma célula-guarda estomática, um tricoma, um grão de pólen em germinação e uma célula do xilema possuem arquiteturas muito diferentes. Essas diferenças não se explicam apenas por genes expressos ou organelas presentes; dependem de como a célula organiza seu interior e sua superfície. O citoesqueleto traduz programas genéticos e sinais ambientais em forma, polaridade e movimento. Ele é uma ponte entre informação molecular e morfologia celular.
No contexto geral da organização eucarionte, o citoesqueleto ocupa um papel integrador comparável ao do sistema de endomembranas, mas em outra dimensão. O sistema de endomembranas organiza fluxos de membranas e moléculas; o citoesqueleto organiza espaço, força, direção e movimento. Juntos, eles permitem que organelas sejam posicionadas, vesículas sejam transportadas, células mudem de forma, membranas sejam remodeladas e divisões celulares sejam executadas. A compartimentalização eucariótica seria incompleta sem uma rede capaz de ordenar os compartimentos no espaço e conectá-los mecanicamente. A célula eucarionte não é apenas compartimentalizada; é arquitetada.
Essa arquitetura dinâmica prepara a compreensão dos capítulos seguintes porque atravessa praticamente todos os processos celulares. O tráfego vesicular depende dele; mitocôndrias, cloroplastos e peroxissomos interagem com ele; o núcleo é posicionado e mecanicamente conectado por ele; a divisão celular exige sua reorganização profunda; a comunicação celular frequentemente modifica sua dinâmica; a forma de tecidos depende de sua atividade. Mesmo quando o citoesqueleto não é o protagonista aparente de um processo, ele frequentemente fornece a infraestrutura física que permite ao processo ocorrer. A célula viva é química, genética e energética, mas também é mecânica. O citoesqueleto é a expressão mais clara dessa dimensão mecânica integrada à vida molecular.
6.8 — O núcleo como centro de armazenamento, regulação e integração genética #
Entre todos os compartimentos da célula eucarionte, o núcleo ocupa posição singular porque concentra a maior parte da informação genética celular e organiza os processos que determinam quando, onde e em que intensidade essa informação será utilizada. A presença de um núcleo delimitado por membrana é uma das características definidoras da célula eucarionte, mas sua importância não se resume a separar o DNA do citosol. O núcleo permite que o material genético seja compactado, protegido, replicado, reparado, transcrito e regulado em um ambiente próprio, ao mesmo tempo em que mantém comunicação seletiva com o restante da célula. Essa separação espacial mudou profundamente a lógica da expressão gênica: nas células procariontes, transcrição e tradução podem ocorrer de modo acoplado no mesmo compartimento; nas células eucariontes, a transcrição ocorre no núcleo, os RNAs são processados antes de sua exportação, e a tradução acontece no citosol ou associada ao retículo endoplasmático. A célula ganha, com isso, novas camadas de controle. [Figura 6.29]
O núcleo é delimitado pelo envoltório nuclear, uma estrutura formada por duas membranas concêntricas separadas por um espaço perinuclear. A membrana externa é contínua com o retículo endoplasmático, o que reforça a integração estrutural entre núcleo e sistema de endomembranas. A membrana interna, por sua vez, associa-se a proteínas específicas e à lâmina nuclear em muitas células animais, contribuindo para a sustentação do núcleo, a organização da cromatina e a manutenção da forma nuclear. Essa arquitetura permite que o núcleo seja simultaneamente protegido e conectado. Ele não é uma estrutura isolada no citoplasma, mas um compartimento integrado à rede membranosa, ao citoesqueleto, ao tráfego molecular e aos sinais celulares.
A comunicação entre núcleo e citoplasma ocorre por meio dos complexos de poro nuclear. Esses poros não são simples aberturas passivas. São grandes estruturas proteicas que atravessam o envoltório nuclear e regulam a passagem de moléculas entre os dois compartimentos. Pequenas moléculas podem atravessar com relativa facilidade, mas proteínas maiores, RNAs e complexos ribonucleoproteicos dependem de mecanismos seletivos de importação e exportação. Proteínas destinadas ao núcleo carregam sinais de localização nuclear reconhecidos por receptores de transporte; RNAs mensageiros processados são exportados em associação com proteínas específicas; subunidades ribossômicas montadas no nucléolo atravessam os poros em direção ao citoplasma. Essa comunicação seletiva é indispensável porque o núcleo precisa receber proteínas regulatórias, enzimas, histonas e fatores de transcrição, ao mesmo tempo em que exporta RNAs e componentes necessários à síntese proteica. [Figura 6.30]
A existência de poros nucleares mostra que a compartimentalização eucariótica não significa isolamento absoluto. O núcleo separa processos, mas não interrompe fluxos. Pelo contrário, sua função depende de uma troca intensa e controlada com o citoplasma. Proteínas sintetizadas no citosol entram no núcleo para participar da replicação do DNA, do reparo, da transcrição, do processamento de RNA e da organização cromatínica. RNAs produzidos no núcleo deixam esse compartimento para serem traduzidos ou exercer funções regulatórias no citoplasma. Sinais vindos da membrana plasmática, de organelas, do estado metabólico ou de estímulos ambientais podem alcançar o núcleo por meio de cascatas de sinalização que modificam a atividade de fatores de transcrição. O núcleo, portanto, funciona como um centro de decisão molecular sensível ao estado global da célula.
O DNA nuclear não se encontra livre e desorganizado. Ele está associado a proteínas, principalmente histonas, formando a cromatina. Essa organização resolve um problema físico evidente: moléculas de DNA extremamente longas precisam ser acomodadas em um compartimento microscópico sem perder completamente a capacidade de serem acessadas por enzimas e proteínas regulatórias. A compactação cromatínica permite armazenar grandes genomas, mas também cria níveis de controle sobre o acesso à informação genética. Regiões mais compactadas tendem a ser menos acessíveis à transcrição, enquanto regiões mais abertas permitem maior interação com fatores regulatórios e RNA polimerases. A cromatina, portanto, não é apenas embalagem do DNA; é parte ativa da regulação gênica. [Tabela 6.8]
Essa relação entre compactação e função é essencial para compreender a célula eucarionte. O mesmo genoma pode produzir células muito diferentes porque diferentes conjuntos de genes são ativados ou silenciados em cada contexto. Uma célula muscular, uma célula epitelial, um neurônio, uma célula do mesófilo foliar, uma célula-guarda estomática e uma célula meristemática podem conter essencialmente a mesma informação genética nuclear dentro de um organismo, mas expressam partes distintas dessa informação. A identidade celular não depende apenas dos genes existentes, mas dos genes que estão acessíveis, ativos, reprimidos, modulados ou preparados para responder a sinais. O núcleo é o compartimento onde essa seleção funcional da informação se organiza.
A cromatina também possui organização espacial dentro do núcleo. Cromossomos não ocupam posições completamente aleatórias, e regiões específicas do genoma podem associar-se a domínios nucleares relacionados à atividade transcricional, repressão, replicação ou reparo. Essa organização tridimensional influencia a probabilidade de interação entre regiões regulatórias e genes, a coordenação de programas transcricionais e a estabilidade do genoma. A célula eucarionte, portanto, organiza a informação genética não apenas em sequência linear de nucleotídeos, mas em arquitetura nuclear. O genoma funciona no espaço. A posição de uma região cromossômica, seu estado de compactação, suas modificações químicas e suas interações com proteínas nucleares contribuem para determinar sua atividade.
O nucléolo é uma das estruturas mais evidentes dentro do núcleo e exemplifica como a organização nuclear pode ocorrer sem membrana delimitante. Ele se forma em torno de regiões cromossômicas que contêm genes de RNA ribossômico e concentra a transcrição desses genes, o processamento de rRNAs e a montagem inicial das subunidades ribossômicas. O nucléolo não é cercado por bicamada lipídica, mas mantém identidade própria por concentração de moléculas, interações dinâmicas e organização funcional. Essa característica reforça uma ideia já introduzida ao discutir o citosol: a compartimentalização celular nem sempre depende de membranas. Dentro do núcleo, corpos nucleares e domínios funcionais permitem concentrar processos específicos sem interromper a continuidade do nucleoplasma. [Figura 6.31]
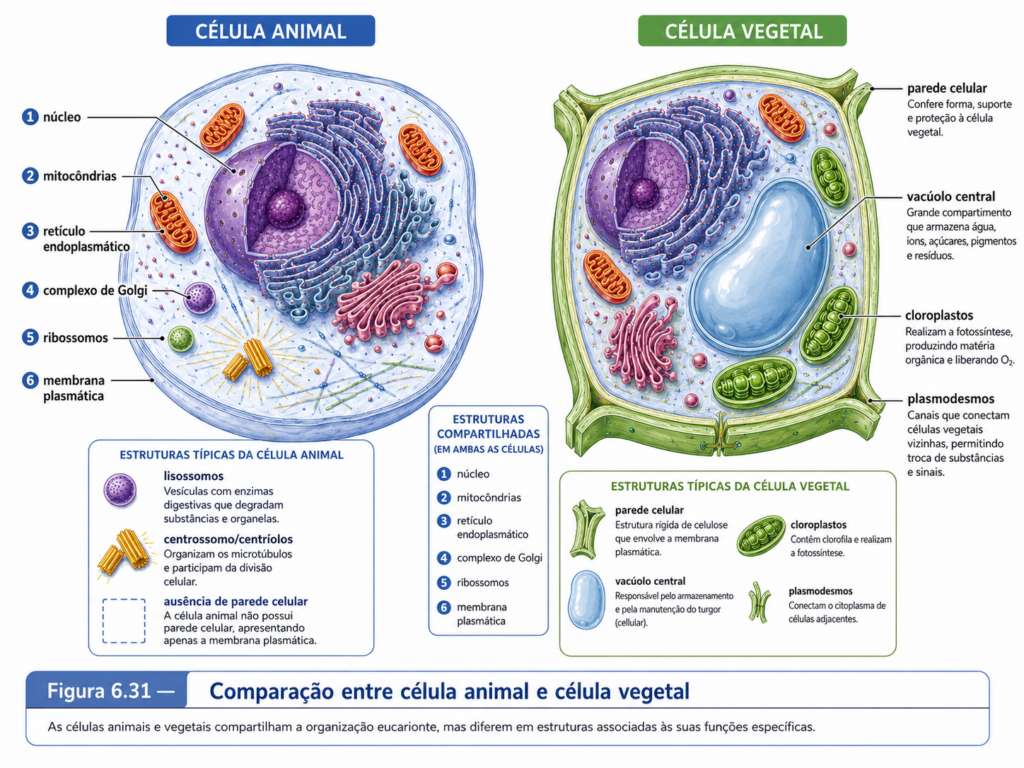
A produção de ribossomos revela a intensa cooperação entre núcleo e citoplasma. Os genes ribossômicos são transcritos no nucléolo; os RNAs ribossômicos são processados e associados a proteínas ribossômicas que foram sintetizadas no citosol e importadas para o núcleo; as subunidades ribossômicas em formação são exportadas pelos poros nucleares; no citoplasma, completarão sua maturação e participarão da tradução de mRNAs. Esse fluxo mostra que mesmo uma estrutura nuclear aparentemente interna, como o nucléolo, depende de proteínas produzidas fora do núcleo e envia produtos que só terão função plena no citoplasma. A separação nuclear cria controle, mas a função celular exige circulação.
A separação entre transcrição e tradução é uma das grandes consequências da origem do núcleo. Em células procariontes, um mRNA pode começar a ser traduzido antes mesmo de sua transcrição ser concluída. Em células eucariontes, o RNA recém-transcrito passa por etapas de processamento antes de atingir o citoplasma. O pré-mRNA pode receber cap na extremidade 5’, cauda poli-A na extremidade 3’ e sofrer remoção de íntrons por splicing. Esses processos influenciam estabilidade, exportação, localização e tradução do mRNA. Além disso, o splicing alternativo permite que um mesmo gene contribua para a produção de diferentes isoformas proteicas, ampliando a complexidade funcional do genoma. O núcleo, nesse sentido, não apenas copia informação do DNA para RNA; ele processa e edita o potencial informacional dos transcritos.
Essa capacidade regulatória ajuda a explicar por que organismos eucarióticos podem gerar grande diversidade celular sem depender exclusivamente de aumento proporcional no número de genes. A complexidade surge também do modo como genes são regulados, combinados, processados e integrados em redes. Fatores de transcrição, modificações da cromatina, RNAs regulatórios, processamento alternativo e controle de exportação nuclear adicionam níveis de decisão entre DNA e proteína. A informação genética não flui como uma linha reta simples; ela passa por filtros, permissões, bloqueios, ajustes e combinações. O núcleo é o principal local onde essas camadas iniciais de regulação são aplicadas.
Em células vegetais, essa regulação nuclear é decisiva para a resposta ao ambiente. Plantas não podem deslocar-se para evitar seca, salinidade, excesso de luz, deficiência nutricional, patógenos ou temperaturas extremas. Sua sobrevivência depende da capacidade de perceber mudanças e reprogramar metabolismo, crescimento e desenvolvimento. Muitos desses ajustes chegam ao núcleo na forma de sinais hormonais, alterações redox, variações de cálcio, mudanças metabólicas ou sinais vindos de cloroplastos, mitocôndrias e peroxissomos. Em resposta, genes relacionados a defesa, ajuste osmótico, proteção antioxidante, remodelamento de parede celular, fechamento estomático, síntese de proteínas protetoras e alteração do crescimento podem ser ativados ou reprimidos. A adaptação da planta ao ambiente passa, portanto, pela capacidade do núcleo de integrar sinais e modificar programas de expressão gênica. [Quadro 6.9]
Essa integração é especialmente importante em condições do semiárido, onde a célula vegetal frequentemente precisa responder a déficits hídricos, alta irradiância e temperaturas elevadas. O cloroplasto pode sinalizar excesso de energia luminosa ou desequilíbrio redox; a membrana plasmática pode perceber alterações osmóticas; o vacúolo participa do ajuste de solutos; mitocôndrias e peroxissomos ajustam metabolismo oxidativo; o citosol transmite sinais por Ca²⁺, fosforilação e metabólitos; o núcleo reorganiza a expressão gênica. A resposta fisiológica observada em uma planta — maior tolerância à seca, manutenção parcial da fotossíntese, redução de crescimento, acúmulo de osmoprotetores ou ativação de defesas — nasce dessa rede. O núcleo não é apenas depósito de DNA; é um centro integrador que converte sinais celulares em mudanças de programa biológico.
A comunicação entre organelas e núcleo é particularmente relevante nas células vegetais porque cloroplastos e mitocôndrias possuem genomas próprios, mas dependem amplamente do genoma nuclear. A maior parte das proteínas dessas organelas é codificada no núcleo, traduzida no citosol e importada posteriormente. Assim, para montar fotossistemas, complexos respiratórios, enzimas metabólicas e sistemas antioxidantes, a célula precisa coordenar expressão gênica nuclear com o estado funcional das organelas. Quando um cloroplasto está danificado ou submetido a estresse luminoso, não faria sentido o núcleo continuar produzindo indiscriminadamente proteínas fotossintéticas como se o sistema estivesse em equilíbrio. Sinais retrógrados vindos de cloroplastos e mitocôndrias ajustam a expressão nuclear, permitindo compatibilizar produção de componentes com necessidade real e capacidade funcional da organela. [Figura 6.32]
O núcleo também precisa preservar a estabilidade do genoma. O DNA está sujeito a danos causados por erros de replicação, radiação, espécies reativas de oxigênio, substâncias químicas e tensões mecânicas. A célula eucarionte possui sistemas de reparo que identificam e corrigem diferentes tipos de lesões. A organização nuclear contribui para esses processos ao reunir enzimas, sensores e regiões cromatínicas em arranjos funcionais. Durante a replicação, o genoma precisa ser copiado com alta fidelidade; durante a divisão celular, os cromossomos precisam ser segregados corretamente; durante a vida celular, lesões precisam ser reparadas antes de comprometerem genes essenciais ou causarem instabilidade. O núcleo é, portanto, também um compartimento de vigilância genômica.
A replicação do DNA nuclear impõe uma coordenação precisa entre cromatina, ciclo celular e divisão. Em células eucariontes, o DNA não é replicado continuamente em qualquer momento, mas em uma fase específica do ciclo celular. Esse controle evita duplicações indevidas e garante que a célula só avance para divisão quando o genoma tiver sido copiado de maneira adequada. Em organismos multicelulares, esse controle é essencial para desenvolvimento, manutenção de tecidos e prevenção de proliferação desordenada. Em plantas, a atividade dos meristemas depende da regulação de ciclos celulares, replicação, diferenciação e resposta ambiental. O núcleo, nesse contexto, conecta informação genética a decisões de crescimento e divisão.
A estrutura nuclear também muda durante o ciclo celular. Em muitas células eucarióticas, o envoltório nuclear se desorganiza durante a mitose e se reorganiza ao redor dos cromossomos segregados ao final da divisão. A cromatina, que durante a interfase pode apresentar regiões mais ou menos compactadas, condensa-se em cromossomos visíveis durante a mitose, facilitando sua distribuição para células-filhas. Essa alternância entre um estado mais funcionalmente acessível e um estado altamente compactado revela a plasticidade da organização nuclear. O núcleo precisa ser, em momentos diferentes, um centro ativo de expressão gênica e uma estrutura capaz de embalar e distribuir cromossomos com precisão. [Animação 6.5]
Em células vegetais, algumas particularidades da divisão reforçam a integração entre núcleo, citoesqueleto e sistema de endomembranas. A formação da placa celular, que dará origem à nova parede entre células-filhas, depende do direcionamento de vesículas para a região central da célula em divisão. Ao mesmo tempo, os cromossomos precisam ser separados adequadamente pelo fuso mitótico. O núcleo organiza o material genético, mas sua divisão efetiva depende de microtúbulos, motores moleculares, vesículas, membranas e síntese de parede. A herança genética e a construção física de novas células ocorrem como eventos coordenados, não como processos independentes.
A forma e a posição do núcleo também têm significado funcional. Em muitas células, o núcleo ocupa posição característica e pode deslocar-se durante desenvolvimento, migração, diferenciação ou resposta a estímulos. Em células vegetais altamente vacuoladas, o núcleo pode ficar deslocado para a periferia, suspenso em cordões citoplasmáticos ou reposicionado conforme o estado celular. Essa posição depende de interações com o citoesqueleto e com a organização do citoplasma. Em células animais, conexões entre citoesqueleto e lâmina nuclear permitem transmissão de forças até o núcleo, influenciando forma nuclear, organização da cromatina e, em alguns contextos, expressão gênica. A informação genética não está isolada das propriedades físicas da célula; ela é armazenada em um compartimento mecanicamente integrado ao citoplasma.
O núcleo também participa da diferenciação celular ao estabilizar programas de expressão gênica. Durante o desenvolvimento, células inicialmente semelhantes passam a assumir identidades distintas. Parte dessa diferenciação envolve ativação de certos genes e repressão de outros, acompanhada por mudanças duradouras na cromatina. Modificações de histonas, metilação do DNA e reorganização de domínios cromatínicos podem contribuir para manter padrões de expressão ao longo de divisões celulares. Essa memória celular permite que uma célula diferenciada preserve sua identidade mesmo após ciclos de divisão. Ao mesmo tempo, determinadas condições experimentais podem reprogramar células, mostrando que a identidade nuclear é estável, mas não absolutamente irreversível. Essa plasticidade tem implicações profundas para desenvolvimento, regeneração, biotecnologia e medicina.
Em plantas, a plasticidade nuclear e celular é particularmente expressiva. Células vegetais diferenciadas podem, em muitos contextos, readquirir competência proliferativa, formar calos, regenerar órgãos ou originar plantas inteiras em condições adequadas de cultura de tecidos. Essa capacidade depende de reprogramação da expressão gênica, resposta hormonal, reorganização cromatínica, divisão celular e diferenciação. A cultura de tecidos vegetais, usada em micropropagação, conservação, transformação genética e melhoramento, explora diretamente essa plasticidade. O núcleo, nesse caso, não é apenas um arquivo de informação herdada; é o local onde programas celulares podem ser redirecionados sob estímulos hormonais e ambientais específicos. [Quadro 6.10]
A biotecnologia moderna depende intensamente da compreensão das funções nucleares. A transformação genética, a edição gênica, a regulação de promotores, a expressão de proteínas recombinantes, a produção de plantas transgênicas e o uso de ferramentas como CRISPR/Cas exigem acesso à informação nuclear e capacidade de modificar sua expressão ou sequência. No entanto, alterar o DNA não garante automaticamente o resultado desejado. A expressão de um gene introduzido depende de sua inserção, do contexto cromatínico, do promotor utilizado, da estabilidade do RNA, do processamento, da tradução e do destino da proteína produzida. A biotecnologia eficaz precisa considerar o núcleo como ambiente regulado, não como um simples local onde genes são depositados.
A organização nuclear também impõe desafios à interpretação de genomas. Sequenciar DNA revela a ordem de nucleotídeos, mas não mostra sozinho quando um gene será ativado, em qual tecido, sob qual condição ambiental, em qual estágio de desenvolvimento ou com qual intensidade. Para compreender a função genética, é necessário integrar genômica, transcriptômica, epigenética, proteômica, fisiologia e biologia celular. O núcleo é o local onde muitas dessas dimensões se cruzam: sequência, cromatina, transcrição, processamento de RNA e resposta a sinais. A leitura do genoma depende do estado celular.
A separação nuclear também contribuiu para a evolução da complexidade eucariótica. Ao desacoplar transcrição e tradução, a célula pôde introduzir etapas intermediárias de controle, como processamento de RNA, exportação seletiva e regulação pós-transcricional. Ao organizar o DNA em cromatina, pôde compactar genomas maiores e controlar acessibilidade. Ao desenvolver poros nucleares, pôde regular o trânsito entre informação genética e execução citoplasmática. Ao integrar sinais vindos de organelas, membranas e metabolismo, pôde ajustar expressão gênica a estados internos e externos. O núcleo, portanto, não representa apenas uma “caixa” para o DNA; representa uma inovação organizacional que ampliou a capacidade regulatória da célula.
Essa inovação tem custo. Manter um núcleo exige energia, proteínas estruturais, sistemas de transporte, controle de montagem e desmontagem durante divisão, organização cromatínica, reparo e vigilância molecular. O envoltório nuclear precisa ser preservado; os poros precisam funcionar; RNAs precisam ser processados e exportados; proteínas precisam ser importadas; cromossomos precisam ser replicados e segregados. A vantagem eucariótica não está na simplicidade, mas na capacidade de sustentar essa complexidade com controle. O núcleo adiciona etapas, mas essas etapas permitem decisões mais refinadas.
Em organismos multicelulares, essa capacidade regulatória torna-se indispensável. As células de um mesmo organismo compartilham origem comum e informação genética, mas precisam exercer funções diferentes. Um tecido vegetal condutor, um meristema, uma epiderme foliar, uma célula-guarda, uma célula do parênquima clorofiliano e uma célula de raiz apresentam programas distintos de expressão gênica. Em animais, neurônios, hepatócitos, células musculares e leucócitos também expressam subconjuntos diferentes do genoma. A multicelularidade depende dessa diferenciação coordenada. O núcleo permite que a mesma informação seja interpretada de modos distintos conforme sinais, posição, história celular e ambiente.
A relação entre núcleo e ambiente é particularmente relevante para organismos sésseis como plantas. Luz, temperatura, disponibilidade de água, nutrientes, patógenos, herbivoria, gravidade e contato físico influenciam programas nucleares. Fotorreceptores podem modificar expressão de genes relacionados a desenvolvimento e fotossíntese; hormônios como auxina, citocinina, ácido abscísico, etileno, giberelinas e jasmonatos modulam redes transcricionais; sinais de estresse alteram fatores de transcrição e cromatina; patógenos ativam genes de defesa. A forma final da planta resulta de uma conversa permanente entre genoma e ambiente, mediada por sinais celulares que convergem no núcleo.
Ao mesmo tempo, o núcleo depende das demais organelas para executar suas decisões. Um gene nuclear pode codificar uma proteína de cloroplasto, mas essa proteína precisará ser traduzida no citosol e importada para o plastídio. Um fator de transcrição pode induzir enzimas antioxidantes, mas sua efetividade dependerá de cofatores, localização e estado redox. A ativação de genes de parede celular precisará ser acompanhada por síntese, processamento e secreção de componentes pelo sistema de endomembranas. A decisão nuclear só se torna função celular quando é executada por ribossomos, organelas, membranas, citoesqueleto e metabolismo. O núcleo coordena, mas não realiza sozinho a vida celular.
Essa perspectiva evita uma visão geneticamente reducionista. O DNA é fundamental, mas não age fora de contexto. O genoma precisa de uma célula para ser lido, protegido, interpretado e convertido em função. A célula, por sua vez, precisa do genoma para manter, renovar e regular sua organização. Núcleo e citoplasma formam uma unidade funcional. O material genético não paira acima da célula como um programa abstrato; ele está inserido em um compartimento físico, regulado por proteínas, influenciado por sinais e dependente de energia. A célula eucarionte é, ao mesmo tempo, expressão do genoma e condição de funcionamento do genoma.
A compreensão do núcleo como centro de armazenamento, regulação e integração genética prepara a análise mais detalhada de sua estrutura em capítulos posteriores. Aqui, o essencial é reconhecer sua posição na organização eucarionte. O núcleo delimita um espaço próprio para o DNA, organiza cromatina, produz e processa RNAs, monta subunidades ribossômicas, controla a passagem de moléculas por poros nucleares, responde a sinais citoplasmáticos e coordena programas de expressão gênica. Ele não é apenas o compartimento que contém os cromossomos; é o local onde informação hereditária, estado celular, ambiente e desenvolvimento começam a ser integrados em decisões moleculares. Na célula eucarionte, a vida depende da cooperação entre compartimentos, e o núcleo é o compartimento que transforma informação genética em possibilidade regulada de forma, função e resposta.
6.9 — Integração entre organelas: a célula como rede funcional #
A compartimentalização da célula eucarionte só adquire significado pleno quando se reconhece que seus compartimentos operam de forma coordenada. Organelas não são unidades autônomas; constituem nós de uma rede funcional na qual moléculas, energia e sinais circulam continuamente. A eficiência celular depende menos da atividade isolada de cada organela e mais da capacidade de integrar fluxos entre elas. A célula eucarionte funciona como um sistema distribuído, no qual processos bioquímicos são espacialmente organizados, mas funcionalmente interdependentes. [Figura 6.33]
Essa integração ocorre por diferentes mecanismos complementares. O primeiro é o tráfego vesicular, que conecta compartimentos do sistema de endomembranas. Proteínas e lipídios sintetizados no retículo endoplasmático são transportados ao complexo de Golgi, modificados e direcionados a seus destinos finais, incluindo membrana plasmática, lisossomos, vacúolos e secreção extracelular. Esse fluxo não é linear e irreversível. Há tráfego anterógrado e retrógrado, permitindo reciclagem de componentes, manutenção da identidade de compartimentos e correção de desvios. A célula mantém equilíbrio dinâmico entre envio e recuperação de moléculas, garantindo que cada organela preserve sua composição funcional mesmo em constante troca com outras.
O segundo mecanismo envolve contatos físicos entre organelas, que permitem comunicação direta sem necessidade de fusão de membranas. Regiões de proximidade entre retículo endoplasmático e mitocôndrias, por exemplo, facilitam transferência de lipídios e sinalização por cálcio. Em células vegetais, interações entre cloroplastos, peroxissomos e mitocôndrias sustentam rotas metabólicas compartilhadas, como a fotorrespiração. Esses contatos funcionam como plataformas de troca rápida e regulada, reduzindo a dependência exclusiva de vesículas e aumentando a eficiência da integração metabólica. [Figura 6.34]
Os fluxos metabólicos representam a dimensão funcional dessa rede. Em vez de ocorrerem em um único compartimento, muitas vias metabólicas atravessam múltiplas organelas. A fotorrespiração ilustra esse princípio: intermediários gerados no cloroplasto são processados em peroxissomos e mitocôndrias antes de retornarem ao cloroplasto. Nenhuma dessas organelas executa a via completa sozinha. O funcionamento depende da circulação coordenada de metabólitos, da manutenção de gradientes e da sincronização de atividades enzimáticas em compartimentos distintos. Esse tipo de organização permite especialização local sem perda de integração global.
A integração energética segue lógica semelhante. Cloroplastos capturam energia luminosa e produzem compostos orgânicos; mitocôndrias utilizam esses compostos para gerar ATP; peroxissomos participam do controle oxidativo associado a essas reações. O estado redox da célula resulta da interação entre essas organelas, não da atividade isolada de uma delas. Alterações em um componente repercutem nos demais. Um aumento na atividade fotossintética, por exemplo, pode exigir maior capacidade de dissipação de energia e controle de espécies reativas, envolvendo múltiplos compartimentos.
O núcleo ocupa posição central nessa rede ao integrar sinais e coordenar respostas. Informações provenientes do metabolismo, do estado energético, de organelas e do ambiente convergem para mecanismos regulatórios nucleares. A expressão gênica é ajustada em função dessas informações, influenciando a produção de proteínas que atuarão em diferentes compartimentos. Ao mesmo tempo, o núcleo depende do citosol e das organelas para executar suas decisões. Proteínas nucleares são sintetizadas fora do núcleo; RNAs precisam ser exportados; sinais precisam ser transmitidos. A relação é bidirecional: o núcleo coordena, mas também responde.
O citoesqueleto fornece a infraestrutura física que sustenta essa integração. Ele organiza o posicionamento das organelas, orienta o transporte de vesículas e contribui para a polaridade celular. Sem essa rede estrutural, o tráfego intracelular seria desorganizado e ineficiente. O citoesqueleto não apenas conecta pontos no espaço; ele define trajetórias e permite que fluxos ocorram com direção e controle. Em células polarizadas, essa organização é ainda mais evidente, pois o destino das moléculas depende de rotas específicas.
A membrana plasmática integra a célula ao ambiente externo e participa ativamente da rede interna. Processos de endocitose e exocitose conectam o meio extracelular ao sistema de endomembranas. Receptores de membrana detectam sinais externos e iniciam cascatas que alcançam o núcleo e outras organelas. Transportadores e canais regulam entrada e saída de substâncias, influenciando o metabolismo interno. A membrana não é apenas limite; é interface dinâmica entre ambiente e rede celular.
Em células vegetais, essa integração adquire características específicas. O vacúolo central ocupa grande volume e participa do equilíbrio osmótico, armazenamento e degradação, influenciando a distribuição do citosol e das organelas. Cloroplastos, mitocôndrias e peroxissomos formam um eixo metabólico altamente interdependente. A parede celular adiciona uma dimensão estrutural externa que condiciona crescimento e forma. O tráfego vesicular direciona componentes para a parede, enquanto o citoesqueleto orienta sua deposição. A célula vegetal funciona como um sistema integrado em que metabolismo, estrutura e ambiente estão profundamente conectados. [Quadro 6.11]
A integração também se manifesta em respostas a estresses. Em condições de seca, salinidade ou alta irradiância, múltiplos compartimentos são mobilizados. O vacúolo ajusta solutos; a membrana regula transporte iônico; cloroplastos ajustam a fotossíntese; mitocôndrias modulam respiração; peroxissomos controlam espécies reativas; o núcleo altera expressão gênica. A resposta final resulta da coordenação dessas atividades. Nenhuma organela isoladamente determina a adaptação; ela emerge da rede.
A dinâmica temporal é outro aspecto essencial. A integração não ocorre apenas no espaço, mas ao longo do tempo. Sinais rápidos no citosol podem preceder alterações mais lentas na expressão gênica. Mudanças metabólicas podem desencadear reorganização estrutural. A célula sincroniza processos com diferentes escalas temporais, garantindo que respostas sejam eficientes e coerentes. Essa coordenação temporal depende de feedbacks positivos e negativos distribuídos pela rede celular.
A interdependência entre organelas também impõe limites. Alterações em um componente podem desestabilizar o sistema. Falhas no tráfego vesicular podem comprometer múltiplas rotas; disfunções mitocondriais podem afetar energia e sinalização; alterações no retículo endoplasmático podem impactar síntese e dobramento de proteínas. A robustez da célula depende de sua capacidade de compensar essas perturbações, mas essa compensação tem limites. A integração aumenta a eficiência, mas também a sensibilidade a falhas sistêmicas.
A célula eucarionte, portanto, deve ser compreendida como uma rede funcional em que compartimentos especializados estão conectados por fluxos materiais, sinais e estruturas físicas. Essa organização permite alta complexidade e flexibilidade, mas exige coordenação contínua. A função celular emerge da interação entre partes, não da soma de atividades isoladas. A integração entre organelas constitui a base operacional da célula eucarionte e estabelece o fundamento para compreender sua diversidade de formas e funções em organismos distintos.
6.10 — Organização eucarionte em células animais, vegetais e fúngicas #
A arquitetura fundamental da célula eucarionte é compartilhada entre diferentes linhagens, mas sua organização funcional varia de acordo com o contexto evolutivo, fisiológico e ecológico. Células animais, vegetais e fúngicas apresentam o mesmo princípio de compartimentalização interna — núcleo, sistema de endomembranas, organelas energéticas, citoesqueleto e citosol —, porém diferem na composição, na proporção e na integração desses componentes. Essas diferenças não são meramente estruturais; refletem estratégias distintas de aquisição de energia, interação com o ambiente, crescimento e adaptação. [Figura 6.35]
As células animais caracterizam-se pela ausência de parede celular rígida, o que lhes confere maior plasticidade de forma. Essa flexibilidade permite migração, remodelamento da superfície, formação de junções intercelulares complexas e interação dinâmica com a matriz extracelular. A membrana plasmática assume papel central na organização celular, funcionando como interface ativa com o ambiente e com outras células. Organelas como lisossomos são particularmente desenvolvidas, refletindo a importância de processos de digestão intracelular, reciclagem e defesa. Mitocôndrias estão distribuídas de acordo com a demanda energética, podendo ser abundantes em células com alta atividade metabólica. O citoesqueleto apresenta organização complexa, sustentando mobilidade, divisão celular e especialização estrutural. A ausência de plastídios e vacúolos centrais reflete a dependência de fontes externas de energia e nutrientes.
Nas células vegetais, a organização eucarionte incorpora elementos adicionais que redefinem sua dinâmica funcional. A presença de parede celular rígida, composta principalmente por celulose, estabelece uma estrutura externa que confere suporte mecânico, proteção e definição de forma. Essa parede não é estática; é continuamente remodelada por processos que envolvem secreção vesicular, atividade enzimática e orientação do citoesqueleto. O vacúolo central ocupa grande parte do volume celular e desempenha múltiplas funções, incluindo armazenamento de solutos, regulação osmótica, degradação de componentes e manutenção do turgor. Essa organização permite que a célula vegetal cresça por expansão com menor custo de síntese citoplasmática.
Os cloroplastos constituem a principal diferença metabólica das células vegetais em relação às animais. Essas organelas permitem a conversão de energia luminosa em energia química, sustentando a fotossíntese e a produção de compostos orgânicos. A presença de cloroplastos implica uma integração metabólica específica com mitocôndrias e peroxissomos, especialmente em processos como fotorrespiração e controle redox. Além disso, plastídios podem diferenciar-se em formas especializadas, como amiloplastos e cromoplastos, refletindo a diversidade funcional das células vegetais. A célula vegetal, portanto, combina compartimentalização eucarionte com capacidade autotrófica e forte integração metabólica.
As células fúngicas apresentam características intermediárias entre os modelos animal e vegetal, mas com particularidades próprias. Assim como as células vegetais, possuem parede celular, porém composta principalmente por quitina e outros polissacarídeos distintos da celulose. Essa parede confere resistência mecânica e proteção, mas permite crescimento por expansão e formação de estruturas filamentosas, como hifas. Ao contrário das plantas, fungos não possuem cloroplastos e dependem de fontes externas de matéria orgânica, absorvendo nutrientes do ambiente. Vacúolos estão presentes e desempenham funções de armazenamento, degradação e regulação osmótica, semelhantes em parte às células vegetais.
O sistema de endomembranas é bem desenvolvido em células fúngicas, refletindo a necessidade de secreção de enzimas e absorção de nutrientes. Fungos frequentemente liberam enzimas no ambiente para degradar matéria orgânica e, posteriormente, absorvem os produtos dessa degradação. Esse modo de nutrição exige um sistema secretor eficiente e integrado. Mitocôndrias desempenham papel central no metabolismo energético, assim como em células animais. O citoesqueleto organiza o crescimento polarizado das hifas, permitindo expansão direcional e colonização de substratos.
A comparação entre esses três tipos celulares revela que a organização eucarionte é um modelo flexível, capaz de sustentar diferentes estratégias biológicas. A presença ou ausência de parede celular, o tipo de parede, a existência de plastídios, o tamanho e função do vacúolo, a organização do citoesqueleto e a dinâmica do sistema de endomembranas variam conforme as demandas fisiológicas. Essas variações não alteram o princípio básico de compartimentalização, mas demonstram que ele pode ser adaptado a diferentes contextos.
A forma celular está diretamente relacionada a essas diferenças estruturais. Células animais podem assumir formas variadas e mudar rapidamente em resposta a estímulos, enquanto células vegetais tendem a apresentar formas mais definidas pela parede celular. Células fúngicas, especialmente em organismos filamentosos, crescem de maneira polarizada, formando redes de hifas que exploram o ambiente. Essas diferenças de forma refletem modos distintos de interação com o meio, desde mobilidade ativa até crescimento direcionado.
A organização interna também influencia a dinâmica metabólica. Em células vegetais, a integração entre cloroplastos, mitocôndrias e vacúolos permite a produção, armazenamento e redistribuição de energia e metabólitos. Em células animais, a dependência de fontes externas de energia exige sistemas eficientes de captação, processamento e distribuição de nutrientes. Em fungos, a secreção extracelular de enzimas e a absorção subsequente definem um metabolismo voltado para decomposição e assimilação. Cada tipo celular representa uma solução integrada para aquisição e uso de recursos.
A comunicação celular também apresenta diferenças relevantes. Células animais estabelecem junções especializadas que permitem adesão, comunicação direta e coordenação entre células. Em plantas, plasmodesmos conectam citoplasmas de células adjacentes, permitindo troca de moléculas e sinais. Em fungos filamentosos, a continuidade citoplasmática ao longo das hifas permite distribuição de nutrientes e organelas. Essas estratégias refletem diferentes formas de organização multicelular e integração entre células.
A resposta ao ambiente é moldada por essas características estruturais. Plantas, por serem sésseis, dependem fortemente da capacidade de ajustar metabolismo e estrutura celular para enfrentar condições adversas. Animais podem combinar respostas celulares com mobilidade e comportamento. Fungos exploram ambientes por crescimento e secreção de enzimas, adaptando-se a substratos variados. Em todos os casos, a célula eucarionte fornece a base funcional, mas sua organização específica determina a estratégia biológica.
A comparação também evidencia limitações e vantagens de cada modelo. A parede celular vegetal confere resistência, mas limita mobilidade. A ausência de parede em células animais permite flexibilidade, mas exige sistemas adicionais de suporte e proteção. A presença de cloroplastos permite autotrofia, mas requer controle rigoroso de processos fotoquímicos. A nutrição heterotrófica simplifica alguns aspectos metabólicos, mas depende da disponibilidade de recursos externos. Essas diferenças refletem adaptações evolutivas a contextos específicos.
A diversidade de células eucariontes demonstra que a organização básica pode ser modificada sem perder sua lógica central. Núcleo, compartimentalização e integração funcional permanecem como elementos comuns, enquanto componentes adicionais ou modificações estruturais permitem especialização. A célula eucarionte não é um modelo único, mas uma plataforma sobre a qual diferentes estratégias biológicas são construídas.
Essa compreensão é essencial para interpretar fenômenos biológicos e aplicações práticas. Em biotecnologia, a escolha do sistema celular — animal, vegetal ou fúngico — influencia produção de proteínas, metabolismo e resposta a modificações genéticas. Em agricultura, a fisiologia celular vegetal define produtividade e adaptação. Em microbiologia, a organização celular fúngica determina estratégias de crescimento e interação com o ambiente. A célula eucarionte, em suas diferentes formas, fornece a base para essas aplicações.
Ao analisar a organização eucarionte em diferentes contextos, torna-se evidente que a diversidade celular não contradiz a unidade fundamental da célula. Pelo contrário, revela a capacidade desse modelo organizacional de sustentar múltiplas soluções biológicas. A célula eucarionte mantém seus princípios estruturais e funcionais, mas adapta sua organização para atender às demandas de cada linhagem. Essa flexibilidade é uma das razões de seu sucesso evolutivo e de sua relevância em sistemas biológicos complexos.
6.11 — Evolução da complexidade eucariótica #
A organização da célula eucarionte representa uma transição evolutiva de grande magnitude, na qual a simplicidade estrutural relativa das células procariontes deu lugar a um sistema altamente compartimentalizado, integrado e regulado. Essa transformação não ocorreu de forma abrupta, mas como resultado de processos graduais de inovação estrutural, integração metabólica e cooperação entre diferentes componentes celulares. A complexidade eucariótica emerge, portanto, como consequência de múltiplas etapas evolutivas que envolveram tanto modificações internas quanto associações entre organismos distintos. [Figura 6.36]
Um dos eventos centrais nesse processo foi a origem das organelas semiautônomas, particularmente mitocôndrias e cloroplastos. A evidência acumulada indica que essas organelas derivam de ancestrais procariontes que estabeleceram relações simbióticas com uma célula hospedeira. No caso das mitocôndrias, a incorporação de um organismo com capacidade respiratória aeróbia conferiu vantagem energética significativa, permitindo maior eficiência na produção de ATP. Para os cloroplastos, a incorporação de um organismo fotossintético permitiu a exploração da energia luminosa. Esses eventos não devem ser interpretados como simples incorporações, mas como processos de integração progressiva, nos quais os organismos envolvidos perderam autonomia e passaram a funcionar como partes de um sistema unificado.
A integração entre hospedeiro e endossimbionte exigiu reorganização profunda. Genes originalmente presentes nos organismos incorporados foram transferidos para o núcleo da célula hospedeira, enquanto proteínas necessárias ao funcionamento das organelas passaram a ser sintetizadas no citosol e importadas posteriormente. Esse processo reduziu a autonomia genética das organelas, mas aumentou a coordenação celular. A célula eucarionte passou a operar com um sistema genético distribuído, no qual o núcleo exerce controle central sobre componentes originalmente independentes. Essa reorganização genética foi essencial para estabilizar a associação simbiótica e transformá-la em uma estrutura funcional permanente.
A evolução da compartimentalização interna também contribuiu para a complexidade eucariótica. A formação de membranas internas, originando estruturas como o retículo endoplasmático e o complexo de Golgi, permitiu separar processos metabólicos, aumentar eficiência e criar novos níveis de regulação. A invaginação da membrana plasmática e a diferenciação de compartimentos internos ofereceram vantagens funcionais, como a capacidade de sintetizar e processar proteínas de maneira mais controlada, distribuir lipídios e estabelecer rotas de tráfego intracelular. A célula eucarionte passou a organizar suas atividades em regiões especializadas, conectadas por sistemas de transporte e comunicação.
O surgimento do núcleo foi outro marco fundamental. Ao delimitar o material genético em um compartimento próprio, a célula criou condições para separar transcrição e tradução, permitindo maior controle da expressão gênica. Essa separação introduziu etapas intermediárias, como processamento de RNA, que ampliaram a capacidade regulatória. A organização do DNA em cromatina e a existência de poros nucleares reforçaram essa complexidade, possibilitando controle refinado sobre o fluxo de informação genética. O núcleo não apenas protege o genoma, mas estabelece um ambiente regulado para sua utilização.
A evolução do citoesqueleto e de sistemas de transporte intracelular também foi decisiva. Em células maiores e mais complexas, a difusão simples não seria suficiente para garantir distribuição eficiente de moléculas e organelas. A formação de redes de filamentos e a emergência de proteínas motoras permitiram transporte direcionado, posicionamento de organelas e organização espacial. Essa infraestrutura mecânica possibilitou aumento do tamanho celular e diversificação funcional, criando condições para maior complexidade estrutural.
A transição para a complexidade eucariótica envolveu também mudanças na regulação metabólica. A integração entre organelas permitiu a formação de redes metabólicas interdependentes, nas quais diferentes compartimentos contribuem para uma mesma via ou processo. Essa organização favorece eficiência e flexibilidade, mas exige coordenação. A evolução de mecanismos de sinalização e regulação tornou-se essencial para manter o equilíbrio funcional. A célula eucarionte desenvolveu sistemas capazes de integrar informações provenientes de múltiplas fontes e ajustar suas respostas de maneira coordenada.
A complexidade eucariótica trouxe vantagens claras, como maior capacidade de regulação, especialização funcional e adaptação a ambientes variados. No entanto, essas vantagens vieram acompanhadas de custos. A manutenção de organelas, sistemas de transporte, mecanismos de controle e estruturas internas exige energia e recursos. A dependência entre componentes aumenta a sensibilidade a falhas. A célula eucarionte precisa equilibrar esses custos com os benefícios da complexidade, ajustando sua organização conforme as condições ambientais e fisiológicas.
A diversidade de células eucariontes reflete diferentes trajetórias evolutivas a partir de um modelo comum. Linhagens distintas adaptaram a organização básica para atender a demandas específicas. Em plantas, a presença de cloroplastos e parede celular permitiu autotrofia e suporte estrutural. Em animais, a ausência de parede celular favoreceu mobilidade e interação com o ambiente. Em fungos, a combinação de parede celular e nutrição heterotrófica resultou em estratégias próprias de crescimento e absorção. Essas variações mostram que a complexidade eucariótica não é uniforme, mas adaptável.
A evolução da complexidade celular está intimamente ligada à emergência da multicelularidade. A capacidade de diferenciar células, coordenar funções e formar tecidos depende de sistemas regulatórios sofisticados, sustentados pela organização eucarionte. A compartimentalização, a regulação gênica e a integração funcional permitem que células assumam papéis especializados dentro de um organismo. A multicelularidade, por sua vez, amplia a complexidade biológica, criando novos níveis de organização.
A análise evolutiva da célula eucarionte também evidencia a importância da cooperação biológica. A integração entre organismos distintos, como no caso da endossimbiose, demonstra que a evolução não ocorre apenas por competição, mas também por associação e interdependência. A célula eucarionte é resultado de uma fusão funcional de componentes que, em outros contextos, poderiam existir de forma independente. Essa cooperação permitiu explorar novas possibilidades metabólicas e estruturais.
A compreensão da evolução da complexidade eucariótica tem implicações amplas. Ela ajuda a interpretar a organização celular atual, orienta a investigação de processos biológicos e fundamenta aplicações em biotecnologia e medicina. Ao reconhecer que a célula é resultado de processos históricos de integração e adaptação, torna-se possível compreender melhor suas capacidades e limitações.
A célula eucarionte, portanto, não é apenas uma estrutura complexa, mas o resultado de uma trajetória evolutiva que combinou inovação, integração e cooperação. Sua organização reflete compromissos entre eficiência, controle e custo, moldados por pressões ambientais e oportunidades evolutivas. Essa perspectiva histórica complementa a análise estrutural e funcional, oferecendo uma visão mais completa da célula como unidade fundamental da vida.
6.12 — A célula eucarionte como unidade integrada de forma, função e regulação #
A análise da célula eucarionte conduz inevitavelmente a uma síntese: estrutura, função e regulação não constituem dimensões independentes, mas expressões de um mesmo sistema integrado. A forma celular não é um atributo estático; resulta da organização interna, das forças mecânicas, da distribuição de organelas e da interação com o ambiente. A função não é apenas consequência de vias metabólicas isoladas; emerge da coordenação entre compartimentos e fluxos. A regulação, por sua vez, não se limita ao controle gênico, mas envolve comunicação entre organelas, estado metabólico, arquitetura celular e sinais externos. A célula eucarionte só pode ser compreendida plenamente quando essas três dimensões são consideradas simultaneamente. [Figura 6.37]
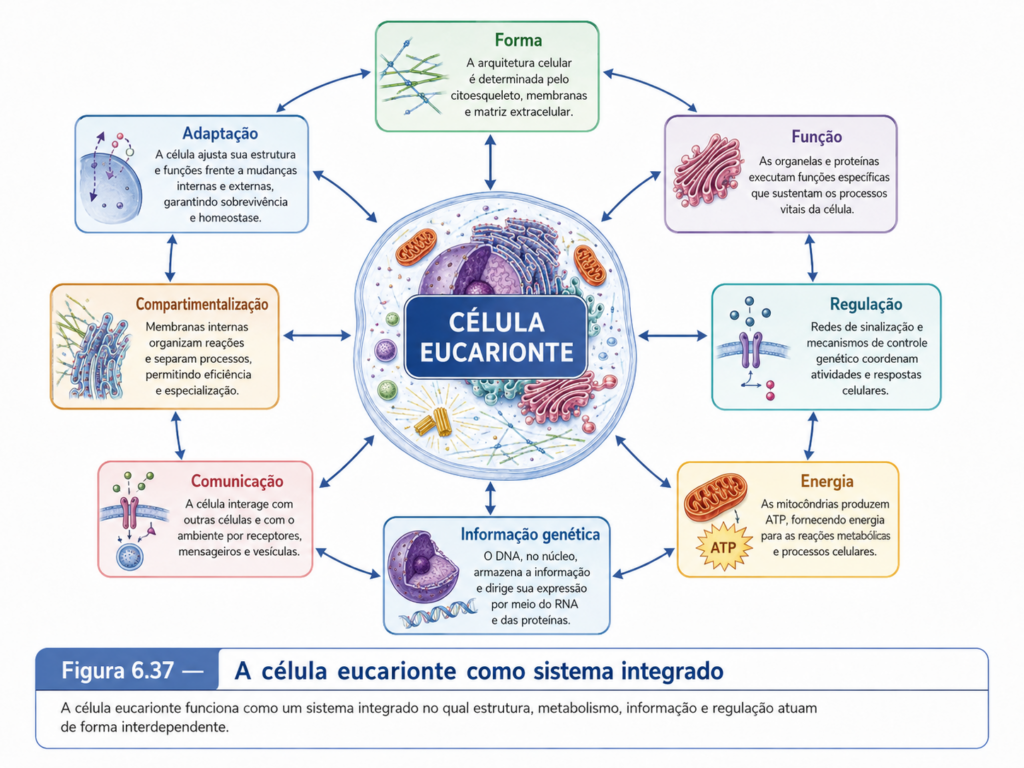
A forma celular reflete diretamente a organização interna. A presença de um grande vacúolo central em células vegetais desloca o citosol para regiões periféricas, condicionando o posicionamento de organelas e o padrão de fluxo citoplasmático. A orientação de microtúbulos influencia a deposição de celulose na parede celular, determinando direções de crescimento. Em células animais, a ausência de parede rígida permite remodelamento contínuo da forma, sustentado pelo citoesqueleto e pela interação com a matriz extracelular. Em fungos, o crescimento polarizado das hifas resulta da organização direcionada do tráfego vesicular e do citoesqueleto. Em todos os casos, a forma não é imposta externamente; é construída a partir da dinâmica interna da célula.
A função celular emerge dessa arquitetura. A fotossíntese depende da organização dos cloroplastos, da distribuição de luz, do estado redox e da interação com mitocôndrias e peroxissomos. A secreção de proteínas depende da integração entre núcleo, retículo endoplasmático, complexo de Golgi, vesículas e membrana plasmática. A respiração celular envolve mitocôndrias, citosol e fluxos metabólicos interconectados. A resposta a estresses ambientais mobiliza múltiplos compartimentos simultaneamente. A célula não executa funções isoladas; ela articula processos distribuídos em uma rede funcional.
A regulação conecta forma e função ao estado do sistema. O núcleo integra sinais e ajusta a expressão gênica, mas sua atividade depende de informações provenientes de organelas e do ambiente. O estado energético, o equilíbrio redox, a disponibilidade de nutrientes e os estímulos externos influenciam diretamente a atividade celular. Cascatas de sinalização, modificações pós-traducionais e reorganização estrutural permitem respostas rápidas e adaptativas. A regulação não atua apenas no início das vias metabólicas; permeia todo o sistema, ajustando continuamente o funcionamento celular.
A integração dessas dimensões torna a célula eucarionte um sistema adaptativo. Em condições estáveis, a célula mantém homeostase, equilibrando síntese e degradação, produção e consumo de energia, estabilidade estrutural e flexibilidade funcional. Em condições de mudança, a célula reorganiza sua arquitetura, redireciona fluxos metabólicos e ajusta programas de expressão gênica. Essa capacidade de adaptação depende da interdependência entre compartimentos e processos. A célula responde como um todo, mesmo quando o estímulo atinge inicialmente apenas uma de suas partes.
A organização integrada também explica a diversidade de formas celulares. Diferentes tipos celulares resultam de variações na expressão gênica, na composição de organelas, na organização do citoesqueleto e na interação com o ambiente. A especialização celular não exige novos componentes fundamentais, mas combinações específicas dos mesmos elementos básicos. A célula eucarionte fornece uma plataforma versátil, capaz de gerar diversidade funcional a partir de uma arquitetura comum.
No contexto vegetal, essa integração assume papel central na adaptação a ambientes variáveis. A célula vegetal precisa coordenar fotossíntese, respiração, armazenamento, transporte e resposta a estresses. A forma da célula, determinada pela parede e pelo vacúolo, influencia a função e a regulação. A resposta a seca, salinidade ou variações térmicas envolve ajustes simultâneos em múltiplos níveis. A produtividade agrícola depende dessa capacidade de integração, pois o desempenho da planta resulta do funcionamento coordenado de suas células.
A biotecnologia e o melhoramento genético também dependem dessa visão integrada. Alterações em genes ou vias metabólicas só produzem efeitos consistentes quando compatíveis com a organização celular. A localização de proteínas, a disponibilidade de cofatores, a interação com outras vias e a capacidade de adaptação do sistema influenciam o resultado. Intervenções eficazes consideram a célula como um sistema, não como um conjunto de partes independentes.
A análise integrada da célula eucarionte reforça a ideia de que a biologia opera em múltiplas escalas. Processos moleculares influenciam estruturas celulares, que por sua vez determinam funções fisiológicas e características de organismos. A célula conecta essas escalas, funcionando como unidade básica de organização da vida. Compreender a célula como sistema integrado permite interpretar fenômenos complexos e aplicar esse conhecimento de forma mais precisa.
A célula eucarionte, portanto, deve ser entendida como uma unidade em que forma, função e regulação são inseparáveis. Sua organização resulta de interações contínuas entre compartimentos, fluxos e sinais, moldadas por pressões evolutivas e ambientais. Essa perspectiva sintetiza os conceitos desenvolvidos ao longo do capítulo e estabelece a base para análises mais detalhadas nos capítulos seguintes, nos quais cada componente será aprofundado sem perder de vista sua inserção no sistema celular como um todo.
Estudo dirigido — Capítulo 6: Organização da Célula Eucarionte #
- Explique por que a compartimentalização é uma característica central da célula eucarionte.
Relacione sua resposta à separação de processos bioquímicos, à formação de microambientes intracelulares e ao aumento da eficiência funcional da célula. - Diferencie citoplasma, citosol e organelas.
Em sua resposta, explique por que o citosol não deve ser entendido como um simples “líquido de preenchimento”, mas como um ambiente molecular organizado e funcional. - Analise a importância do sistema de endomembranas para a rota secretora e endocítica.
Descreva o papel do retículo endoplasmático, do complexo de Golgi, das vesículas, da membrana plasmática, dos lisossomos ou vacúolos na síntese, modificação, transporte, reciclagem e degradação de componentes celulares. - Compare mitocôndrias, cloroplastos e peroxissomos quanto à sua função na célula eucarionte.
Mostre como essas organelas participam da conversão de energia, do metabolismo oxidativo, do controle redox e da integração metabólica, especialmente em células vegetais. - Explique como núcleo, citoesqueleto e organelas atuam de forma integrada na organização da célula eucarionte.
Relacione armazenamento da informação genética, expressão gênica, transporte intracelular, posicionamento de organelas, resposta ao ambiente e manutenção da forma celular.
Questão de pesquisa #
- Pesquise um exemplo de estresse ambiental em plantas, como seca, salinidade ou excesso de luz, e explique como a célula eucarionte vegetal responde a essa condição.
Na resposta, relacione pelo menos três componentes celulares estudados no capítulo, como cloroplastos, mitocôndrias, peroxissomos, vacúolo, núcleo, membrana plasmática ou citoesqueleto. Ao final, cite as fontes consultadas.



