4.1 Estrutura e organização das membranas biológicas #
A delimitação entre o interior da célula e o ambiente externo representa uma condição indispensável para a emergência e manutenção da vida, uma vez que os sistemas biológicos dependem de gradientes químicos e físicos que só podem ser sustentados na presença de uma barreira seletiva. A membrana plasmática surge, nesse contexto, não apenas como um limite estrutural, mas como uma entidade dinâmica, cuja organização molecular permite simultaneamente estabilidade, flexibilidade e interação contínua com o meio. Essa dualidade — barreira e interface — define a natureza das membranas biológicas e orienta sua compreensão em nível molecular.
Do ponto de vista estrutural, as membranas são formadas predominantemente por lipídios organizados em uma bicamada, cuja origem reside nas propriedades anfipáticas dessas moléculas. Fosfolipídios, principais constituintes da membrana, apresentam uma região polar hidrofílica e uma região apolar hidrofóbica, o que determina sua auto-organização espontânea em meio aquoso. Nessa configuração, as cabeças polares orientam-se para os meios intra e extracelular, enquanto as caudas hidrofóbicas se organizam no interior da bicamada, formando uma região central não polar [Figura 4.1]. Essa arquitetura estabelece uma barreira física à difusão livre de moléculas hidrossolúveis, ao mesmo tempo em que permite a passagem seletiva de substâncias lipossolúveis.
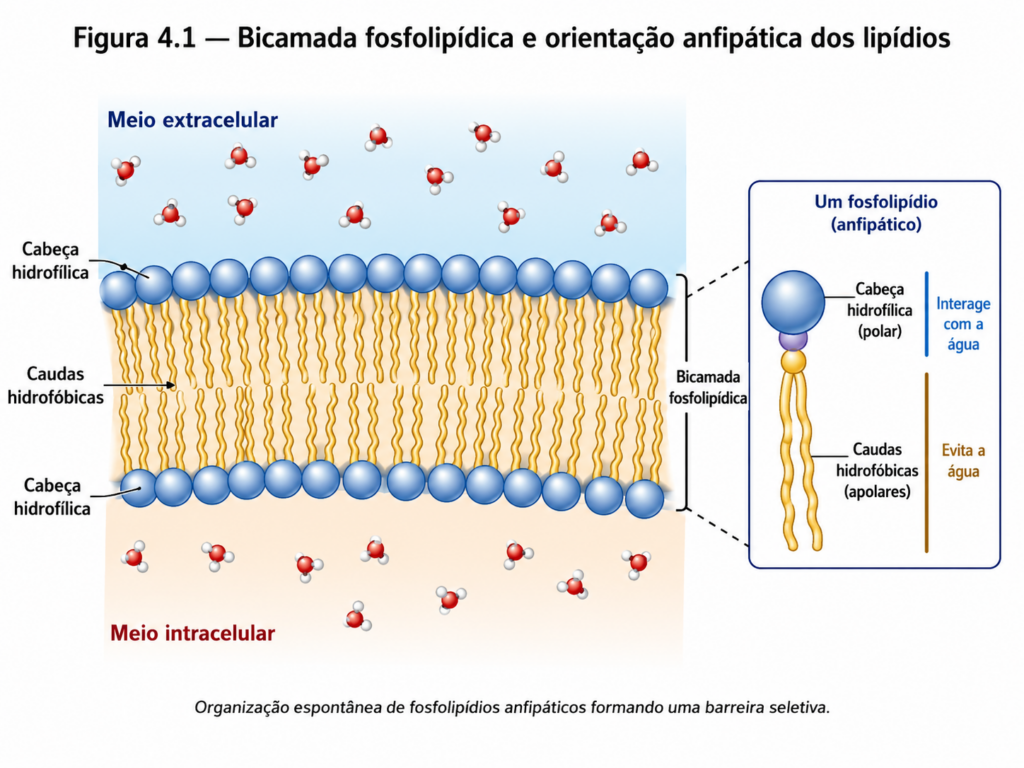
Entretanto, a membrana não é composta exclusivamente por fosfolipídios. Outros lipídios, como colesterol e glicolipídios, desempenham papéis fundamentais na modulação de suas propriedades físico-químicas. O colesterol, por exemplo, atua como regulador da fluidez da membrana, restringindo o movimento excessivo dos fosfolipídios em temperaturas elevadas e impedindo sua compactação em temperaturas mais baixas. Esse efeito estabilizador confere à membrana uma capacidade adaptativa frente a variações ambientais, característica particularmente relevante em organismos submetidos a flutuações térmicas [Figura 4.2].
Além da composição lipídica, um aspecto central da organização das membranas é sua assimetria. As duas faces da bicamada não são idênticas, diferindo tanto na composição lipídica quanto na distribuição de proteínas. Fosfolipídios como fosfatidilserina e fosfatidiletanolamina tendem a localizar-se predominantemente na face citosólica, enquanto glicolipídios e fosfatidilcolina são mais abundantes na face externa. Essa assimetria não é aleatória, mas funcionalmente determinada, estando relacionada a processos como reconhecimento celular, sinalização e manutenção da integridade estrutural [Tabela 4.1].

A presença de proteínas constitui outro elemento essencial na organização das membranas. Essas macromoléculas não apenas se associam à bicamada lipídica, mas participam ativamente de sua funcionalidade, desempenhando papéis que incluem transporte, recepção de sinais, atividade enzimática e ancoragem estrutural. A interação entre lipídios e proteínas não ocorre de forma rígida, mas sim em um ambiente altamente dinâmico, no qual os componentes apresentam mobilidade lateral significativa.
Essa característica dinâmica levou à formulação do modelo do mosaico fluido, que descreve a membrana como uma estrutura bidimensional na qual lipídios e proteínas se organizam de maneira relativamente livre, formando um sistema fluido e heterogêneo. Nesse modelo, as proteínas estão inseridas ou associadas à bicamada lipídica como elementos dispersos em um “mar” lipídico, podendo deslocar-se lateralmente ao longo da membrana [Figura 4.3]. Essa mobilidade não é absoluta, sendo modulada por interações com o citoesqueleto, com outras proteínas e com domínios lipídicos especializados.
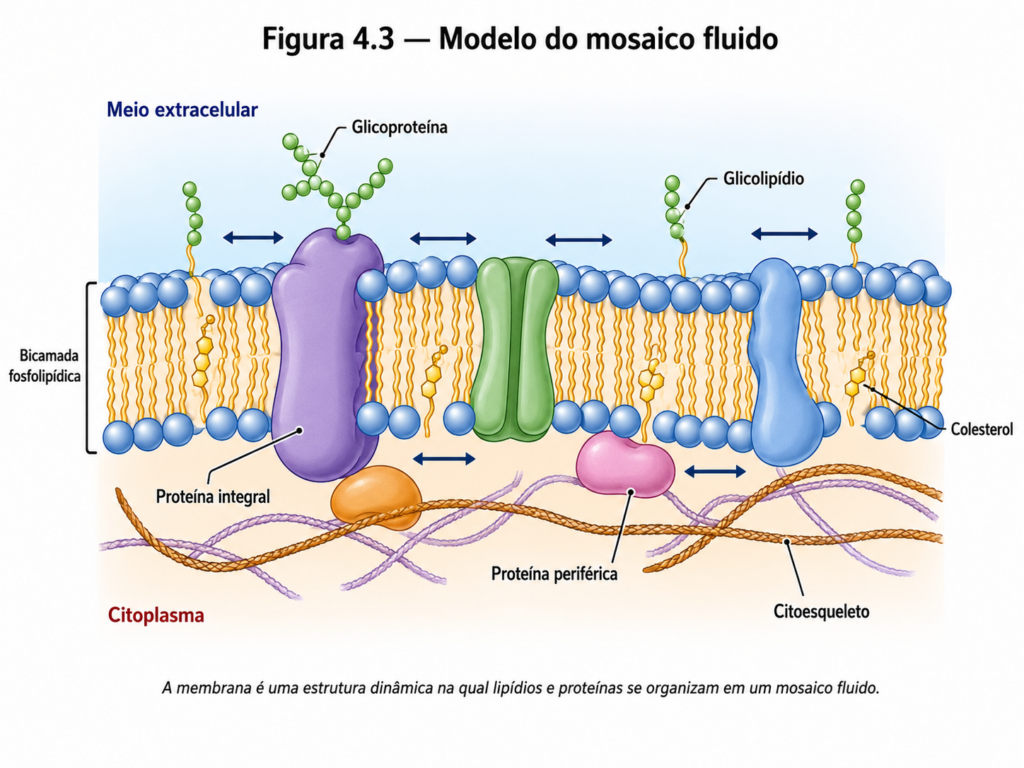
A fluidez da membrana, portanto, não é apenas uma propriedade física, mas um determinante funcional crítico. Ela permite que a membrana responda rapidamente a estímulos, reorganize seus componentes e participe de processos como fusão vesicular, endocitose e formação de complexos de sinalização. A fluidez depende de fatores como o grau de saturação dos ácidos graxos dos fosfolipídios — cadeias insaturadas aumentam a fluidez, enquanto cadeias saturadas promovem maior rigidez — e da presença de colesterol, que atua como modulador estrutural.
Outro aspecto relevante da organização das membranas é a existência de microdomínios lipídicos, frequentemente denominados “rafts”. Esses domínios são regiões enriquecidas em colesterol e esfingolipídios, que apresentam organização mais ordenada em relação ao restante da membrana. Embora sua existência e função ainda sejam objeto de investigação, evidências indicam que esses microdomínios atuam como plataformas para a organização de proteínas envolvidas em sinalização celular e tráfego membranar [Figura 4.4].
A compreensão da estrutura e organização das membranas biológicas exige, portanto, uma visão integrada que considere a interação entre lipídios, proteínas e o ambiente celular. Longe de ser uma barreira estática, a membrana configura-se como um sistema dinâmico e altamente organizado, capaz de ajustar suas propriedades em resposta a condições internas e externas. Essa plasticidade estrutural é o que permite à célula manter sua individualidade ao mesmo tempo em que interage continuamente com o meio, estabelecendo as bases para processos fundamentais que serão explorados nas seções subsequentes.
4.2 Proteínas de membrana: estrutura, classificação e função #
A organização lipídica das membranas estabelece a base estrutural para a compartimentalização celular, mas é a incorporação de proteínas que transforma essa estrutura em um sistema funcional capaz de executar processos biológicos complexos. As proteínas de membrana não apenas ocupam a bicamada lipídica, mas definem a capacidade da célula de interagir com o ambiente, transportar substâncias, perceber sinais e manter sua organização interna. Nesse sentido, a membrana deixa de ser uma barreira passiva e passa a operar como uma plataforma molecular ativa.
As proteínas de membrana podem ser classificadas de acordo com sua forma de associação com a bicamada lipídica. As proteínas integrais estão inseridas na membrana, frequentemente atravessando completamente a bicamada por meio de regiões hidrofóbicas que interagem com o interior lipídico [Figura 4.5]. Essas proteínas transmembrana apresentam domínios estruturais específicos, geralmente organizados em hélices alfa ou, menos frequentemente, em folhas beta, que permitem sua estabilidade em um ambiente hidrofóbico. Já as proteínas periféricas associam-se de maneira mais frouxa à superfície da membrana, interagindo com cabeças polares dos lipídios ou com outras proteínas, sem penetrar no núcleo hidrofóbico da bicamada. Há ainda proteínas ancoradas por lipídios, que se ligam covalentemente a moléculas lipídicas inseridas na membrana, garantindo sua fixação sem a necessidade de regiões transmembrana.
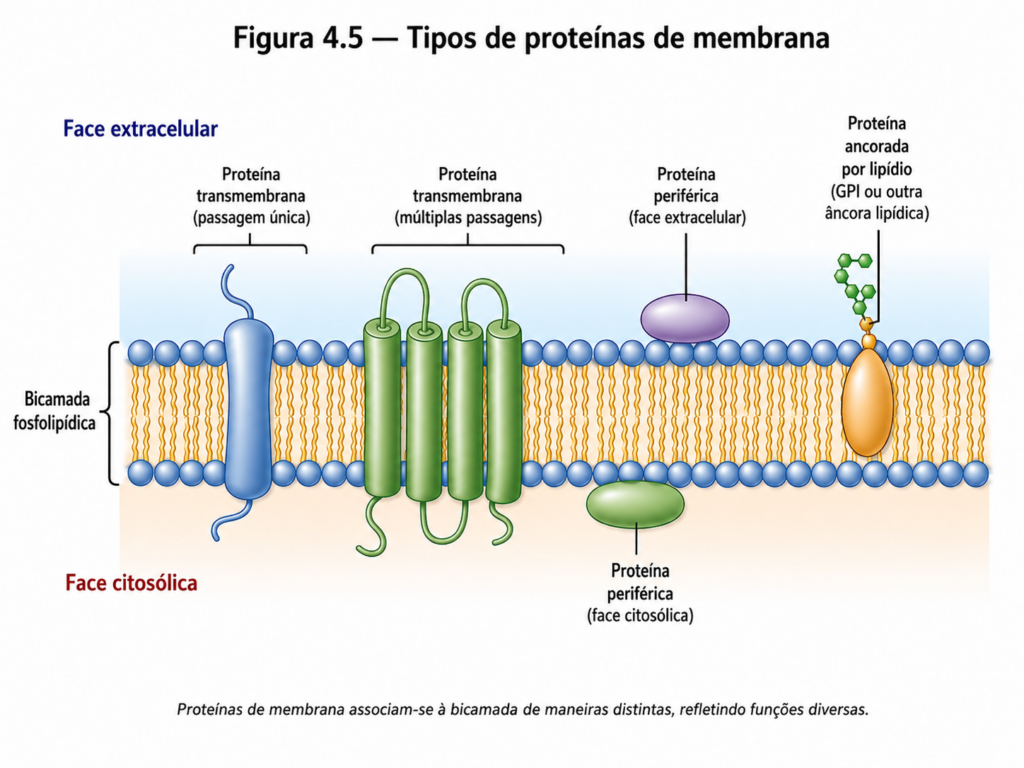
Essa diversidade estrutural reflete diretamente a variedade funcional dessas proteínas. Uma das funções mais críticas é o transporte de moléculas através da membrana, processo inviável apenas pela bicamada lipídica para a maioria das substâncias polares. Proteínas transportadoras e canais iônicos permitem a passagem seletiva de íons e moléculas, estabelecendo fluxos regulados essenciais para a manutenção da homeostase celular [Figura 4.6]. Enquanto canais formam poros hidrofílicos que possibilitam a difusão rápida de íons, proteínas carreadoras operam por meio de mudanças conformacionais que permitem o transporte específico de solutos.
Além do transporte, as proteínas de membrana atuam como receptores, desempenhando papel central na comunicação celular. Esses receptores reconhecem moléculas sinalizadoras, como hormônios, neurotransmissores ou fatores de crescimento, e convertem essa interação em respostas intracelulares. Essa função é particularmente relevante em organismos multicelulares, nos quais a coordenação entre células depende de sistemas eficientes de sinalização [Figura 4.7].
Outro papel fundamental das proteínas de membrana está relacionado à atividade enzimática. Diversas reações bioquímicas ocorrem associadas à membrana, especialmente em processos como metabolismo energético e síntese de biomoléculas. Nessas situações, a organização espacial proporcionada pela membrana favorece a eficiência das reações, permitindo a proximidade entre enzimas e substratos e facilitando a formação de complexos funcionais.
As proteínas também desempenham funções estruturais, ancorando a membrana ao citoesqueleto e à matriz extracelular. Essa ancoragem é essencial para a manutenção da forma celular, para a organização dos tecidos e para a transmissão de forças mecânicas. Em células especializadas, como epitélios, essas interações são determinantes para a formação de domínios funcionais distintos, refletindo a polaridade celular [Figura 4.8].
Apesar dessa organização funcional, as proteínas de membrana não estão fixas em posições estáticas. A maioria apresenta mobilidade lateral, deslocando-se ao longo da bicamada lipídica, o que permite a reorganização dinâmica da membrana em resposta a estímulos. No entanto, essa mobilidade pode ser restrita por interações com o citoesqueleto, com outras proteínas ou com microdomínios lipídicos, resultando em regiões de maior ou menor fluidez e organização.
A distribuição das proteínas na membrana não é aleatória. A célula regula ativamente a localização e a concentração dessas moléculas, criando domínios especializados que concentram funções específicas. Esse padrão de organização é essencial para processos como sinalização, transporte direcionado e interação celular, evidenciando que a membrana funciona como um sistema altamente organizado e funcionalmente compartimentalizado [Tabela 4.2].
A compreensão das proteínas de membrana, portanto, exige uma abordagem integrada que considere sua estrutura, sua inserção na bicamada e sua dinâmica funcional. Elas representam o principal elo entre a organização molecular da membrana e as funções biológicas que dela emergem, estabelecendo as bases para processos que serão aprofundados nas seções seguintes, especialmente aqueles relacionados ao transporte de substâncias e à comunicação celular.
4.3 Propriedades físico-químicas e permeabilidade seletiva #
A existência de uma bicamada lipídica estabelece uma separação física entre o meio intracelular e o ambiente externo, mas é a natureza físico-química dessa estrutura que determina quais substâncias podem atravessá-la e em que condições esse trânsito ocorre. A permeabilidade seletiva das membranas não é uma característica acessória, mas um princípio fundamental que sustenta a organização celular, permitindo a manutenção de gradientes químicos, elétricos e osmóticos indispensáveis ao funcionamento da célula.
Do ponto de vista molecular, a permeabilidade da membrana está diretamente relacionada à composição lipídica da bicamada e às propriedades das moléculas que interagem com ela. O interior hidrofóbico da membrana impõe uma barreira energética significativa à passagem de substâncias polares ou carregadas, enquanto moléculas pequenas e apolares, como gases (O₂ e CO₂), conseguem atravessar a bicamada com relativa facilidade por difusão simples [Figura 4.9]. Esse comportamento decorre da afinidade dessas moléculas pelo ambiente lipídico, permitindo sua dissolução e difusão através da membrana.
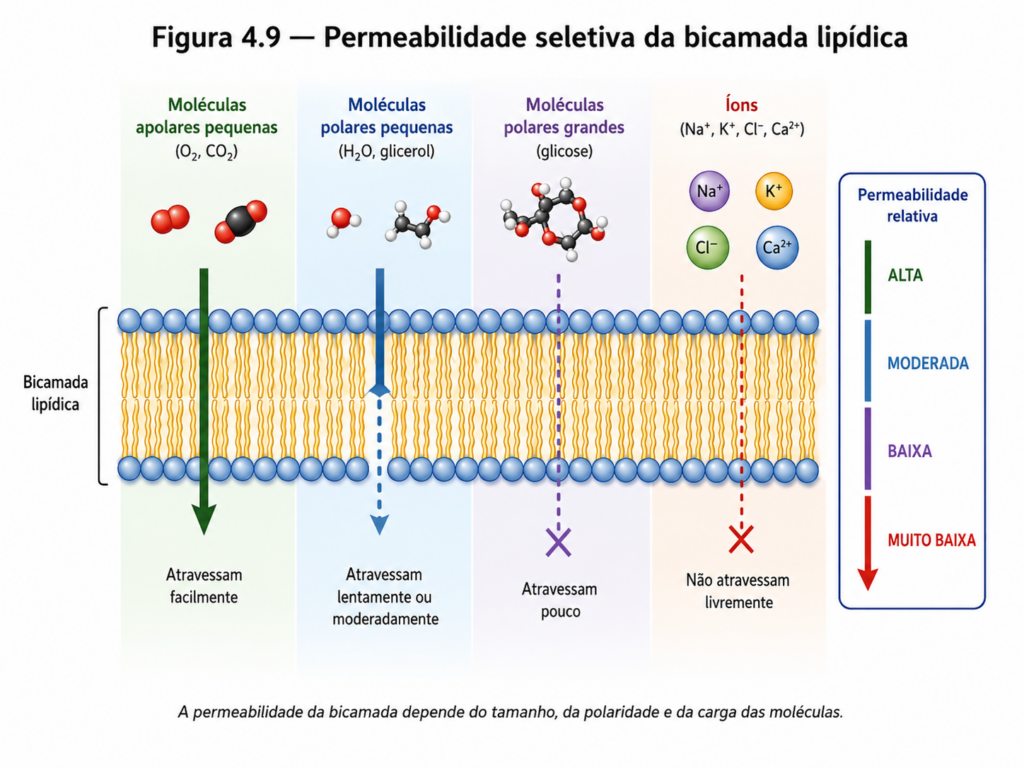
A passagem de moléculas hidrofílicas, por outro lado, é limitada e depende de fatores como tamanho, carga elétrica e grau de hidratação. Moléculas pequenas e polares, como a água, podem atravessar a membrana em taxas moderadas, frequentemente auxiliadas por proteínas específicas, enquanto íons e macromoléculas enfrentam uma barreira praticamente intransponível na ausência de mecanismos especializados. Essa seletividade não apenas restringe o fluxo de substâncias, mas também define a necessidade de sistemas de transporte mediados por proteínas, que serão discutidos nas seções seguintes.
A permeabilidade da membrana pode ser descrita em termos de coeficientes que refletem a facilidade com que diferentes substâncias atravessam a bicamada. Esses coeficientes dependem tanto das propriedades da molécula quanto da organização da membrana, incluindo fatores como fluidez, espessura e composição lipídica. Alterações na proporção de ácidos graxos saturados e insaturados, por exemplo, modificam a fluidez da membrana e, consequentemente, sua permeabilidade, evidenciando a relação direta entre estrutura e função [Tabela 4.3].
Outro elemento central na compreensão da permeabilidade seletiva é a formação de gradientes eletroquímicos. A distribuição desigual de íons entre o interior e o exterior da célula gera diferenças de concentração e de potencial elétrico que influenciam o movimento dessas partículas. O conceito de gradiente eletroquímico integra esses dois componentes — químico e elétrico — e determina a direção e a intensidade do fluxo de íons através da membrana [Figura 4.10]. Esse princípio é essencial para processos como condução nervosa, contração muscular e transporte ativo de solutos.
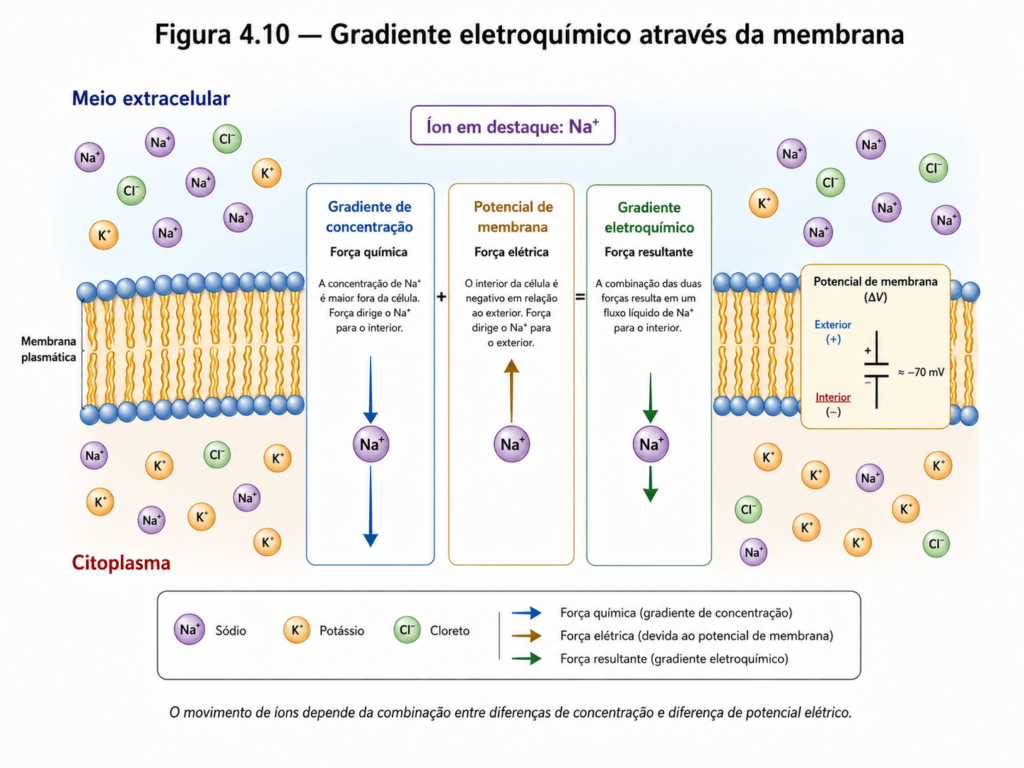
A manutenção desses gradientes depende da baixa permeabilidade da membrana a íons e da ação coordenada de proteínas transportadoras que regulam seu fluxo. A membrana, portanto, não apenas separa compartimentos, mas cria condições para o armazenamento de energia potencial na forma de gradientes, que podem ser utilizados em diversos processos celulares. Essa capacidade de gerar e manter diferenças entre compartimentos é uma das propriedades mais fundamentais dos sistemas vivos.
Além disso, a permeabilidade seletiva contribui para a compartimentalização interna da célula. Organelas delimitadas por membranas apresentam composições químicas distintas, permitindo a ocorrência de reações específicas em ambientes controlados. Essa organização aumenta a eficiência metabólica e possibilita a coexistência de processos que, de outra forma, seriam incompatíveis no mesmo espaço celular [Figura 4.11].
A compreensão das propriedades físico-químicas da membrana e de sua permeabilidade seletiva revela, portanto, que a célula não é um sistema homogêneo, mas um conjunto de compartimentos interdependentes, mantidos por barreiras dinâmicas que regulam o fluxo de matéria e energia. Esse controle seletivo é o que permite à célula sustentar sua organização interna e responder de forma precisa às variações do ambiente, estabelecendo a base para os mecanismos de transporte que serão detalhados nas próximas seções.
4.4 Transporte passivo através da membrana #
A existência de gradientes de concentração entre os compartimentos intra e extracelular cria uma tendência natural ao fluxo de substâncias através da membrana. Esse movimento espontâneo, orientado pela dissipação de energia livre, define o transporte passivo, um conjunto de processos nos quais as moléculas atravessam a membrana sem gasto direto de energia metabólica. Ainda que conceitualmente simples, o transporte passivo assume formas distintas, determinadas pelas propriedades das moléculas e pelas limitações impostas pela bicamada lipídica.
A difusão simples representa a forma mais elementar de transporte passivo. Nesse processo, moléculas atravessam diretamente a bicamada lipídica seguindo seu gradiente de concentração, movendo-se de regiões de maior para menor concentração até o estabelecimento do equilíbrio [Figura 4.12]. Essa modalidade é restrita a substâncias pequenas e apolares ou fracamente polares, como gases e algumas moléculas lipossolúveis, cuja interação com o interior hidrofóbico da membrana permite sua passagem sem a necessidade de intermediários proteicos. A taxa de difusão simples depende de fatores como o gradiente de concentração, a área da membrana e a permeabilidade intrínseca da substância.
Entretanto, a maioria das moléculas biologicamente relevantes não possui características compatíveis com a difusão direta pela bicamada lipídica. Íons, açúcares e aminoácidos, por exemplo, apresentam alta polaridade ou carga elétrica, o que impede sua passagem espontânea. Nesses casos, o transporte passivo ocorre por meio da difusão facilitada, um processo mediado por proteínas específicas que fornecem caminhos hidrofílicos para a travessia da membrana.
As proteínas envolvidas na difusão facilitada podem ser classificadas em dois grupos principais: canais e transportadores. Os canais proteicos formam poros que permitem a passagem rápida de íons ou moléculas pequenas, geralmente de forma altamente seletiva. Muitos desses canais são regulados por mecanismos de abertura e fechamento, podendo responder a estímulos elétricos, químicos ou mecânicos [Figura 4.13]. Essa regulação confere à célula a capacidade de controlar fluxos iônicos de maneira precisa, aspecto fundamental em processos como excitabilidade celular e transmissão de sinais.
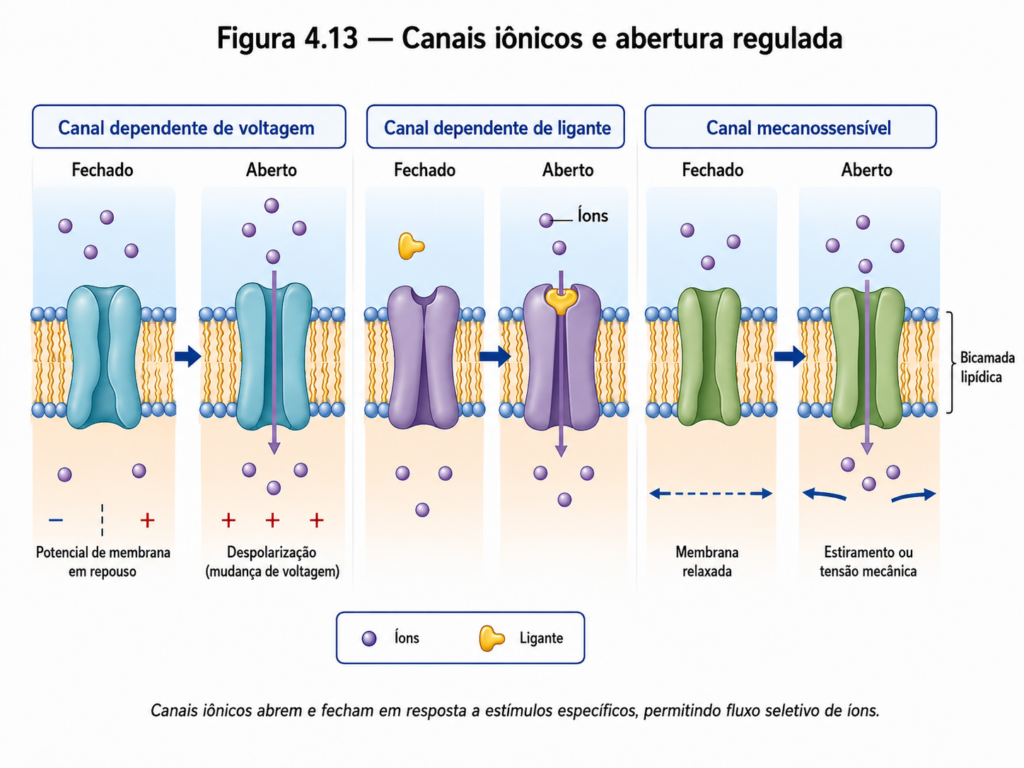
Os transportadores, por sua vez, operam por meio de mudanças conformacionais que permitem o deslocamento de moléculas específicas através da membrana. Diferentemente dos canais, que permitem fluxo contínuo, os transportadores apresentam ciclos de ligação e liberação do soluto, resultando em uma taxa de transporte mais lenta, porém altamente específica. Esse mecanismo implica a existência de um número limitado de sítios de transporte, o que leva a um comportamento de saturação à medida que a concentração do soluto aumenta [Figura 4.14]. Essa característica aproxima a cinética dos transportadores àquela observada em enzimas, evidenciando a natureza altamente regulada desse processo.
Outro aspecto relevante da difusão facilitada é sua especificidade. Cada proteína transportadora reconhece determinados substratos com base em suas propriedades estruturais, garantindo que apenas moléculas específicas sejam transportadas. Essa seletividade é essencial para a manutenção da composição química celular, evitando fluxos indiscriminados que comprometeriam a homeostase.
Apesar de ocorrerem sem gasto direto de ATP, os processos de transporte passivo são profundamente influenciados pelos gradientes eletroquímicos estabelecidos na membrana. A direção do fluxo de íons, por exemplo, depende não apenas da diferença de concentração, mas também do potencial elétrico da membrana, o que pode favorecer ou dificultar o movimento dessas partículas. Assim, o transporte passivo deve ser compreendido como resultado da interação entre forças químicas e elétricas que atuam simultaneamente sobre as moléculas.
A eficiência do transporte passivo também está relacionada à organização da membrana e à distribuição das proteínas transportadoras. Regiões especializadas podem concentrar determinados canais ou transportadores, criando microambientes funcionais que otimizam o fluxo de substâncias conforme as necessidades celulares [Tabela 4.4]. Essa organização espacial reforça a ideia de que a membrana não é homogênea, mas apresenta uma arquitetura funcional complexa.
A análise do transporte passivo evidencia que, mesmo na ausência de gasto energético direto, a célula mantém um controle rigoroso sobre o fluxo de substâncias. Esse controle resulta da combinação entre propriedades físico-químicas da membrana e a ação de proteínas especializadas, garantindo que o movimento de moléculas ocorra de forma seletiva e regulada. Essa base conceitual é essencial para compreender os mecanismos de transporte ativo, nos quais a célula utiliza energia para estabelecer e manter gradientes que sustentam sua organização funcional.
4.5 Transporte ativo e manutenção de gradientes #
A manutenção de diferenças de concentração entre o interior e o exterior da célula constitui uma condição essencial para o funcionamento dos sistemas biológicos, mas tais gradientes não são estáveis por natureza. A tendência termodinâmica favorece sua dissipação, o que implicaria a perda de organização interna e a inviabilização de processos fundamentais. Para contornar essa limitação, a célula utiliza mecanismos de transporte ativo, capazes de deslocar substâncias contra seus gradientes de concentração ou eletroquímicos, à custa de energia metabólica.
O transporte ativo caracteriza-se, portanto, pela necessidade de acoplamento energético, geralmente proveniente da hidrólise de ATP ou da energia armazenada em gradientes previamente estabelecidos. Esse processo permite à célula concentrar íons, nutrientes e outras moléculas em compartimentos específicos, criando condições para o funcionamento de processos como excitabilidade elétrica, síntese de biomoléculas e regulação osmótica [Figura 4.15].
O transporte ativo primário envolve diretamente a utilização de energia química, tipicamente na forma de ATP, para promover mudanças conformacionais em proteínas transportadoras especializadas. Essas proteínas, frequentemente denominadas bombas, possuem sítios específicos de ligação para os solutos e para o ATP, permitindo que a energia liberada na hidrólise seja convertida em trabalho mecânico. Um exemplo clássico é a bomba de sódio e potássio, responsável por exportar íons Na⁺ e importar íons K⁺ contra seus respectivos gradientes, estabelecendo um dos principais gradientes eletroquímicos da célula [Figura 4.16]. Esse gradiente não apenas regula o volume celular, mas também serve como base para diversos processos fisiológicos, incluindo a condução de impulsos nervosos.
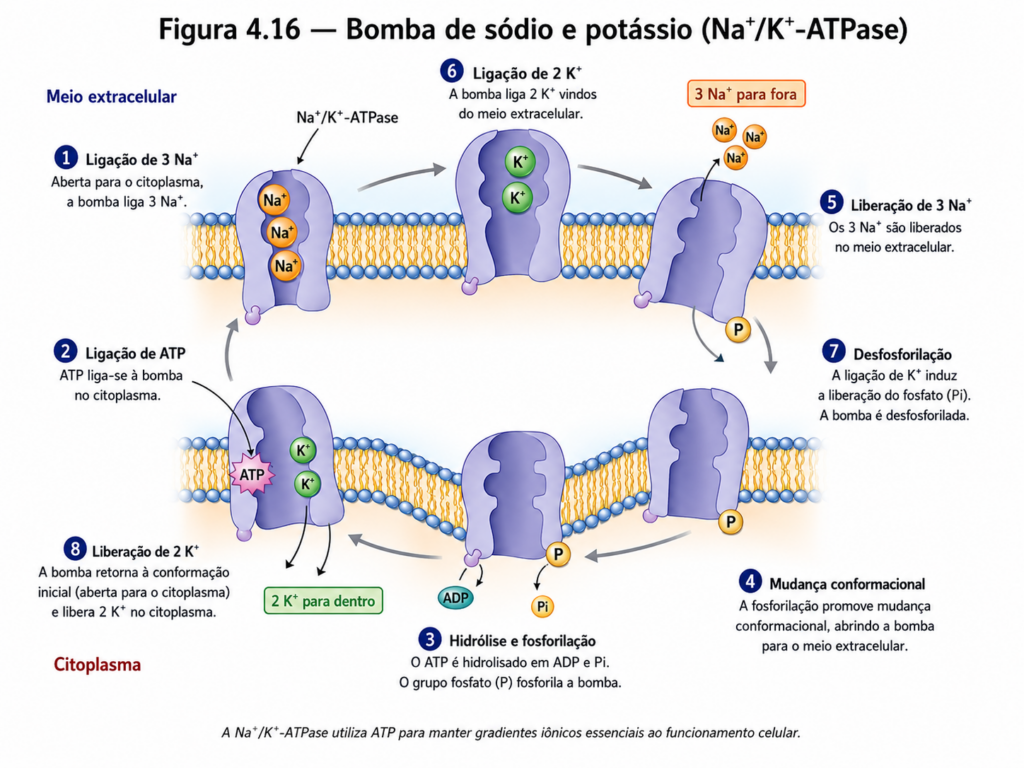
Outras ATPases, como as bombas de cálcio e de prótons, desempenham papéis igualmente críticos, controlando a concentração intracelular de íons que atuam como sinais celulares ou participam de processos metabólicos. A diversidade dessas bombas reflete a necessidade de controle preciso sobre diferentes componentes químicos, evidenciando a centralidade do transporte ativo na homeostase celular [Tabela 4.5].
Em contraste com o transporte ativo primário, o transporte ativo secundário não utiliza diretamente ATP, mas depende da energia armazenada em gradientes eletroquímicos previamente estabelecidos. Nesse caso, o fluxo espontâneo de uma substância a favor de seu gradiente é acoplado ao transporte de outra substância contra o seu gradiente. Esse mecanismo pode ocorrer na forma de simporte, quando ambas as substâncias se deslocam na mesma direção, ou de antiporte, quando se movem em direções opostas [Figura 4.17]. A eficiência desse sistema depende da manutenção dos gradientes que o alimentam, frequentemente gerados por bombas de transporte ativo primário.
O transporte ativo secundário é particularmente relevante na absorção de nutrientes, como glicose e aminoácidos, em células que dependem da captação eficiente de substratos mesmo quando sua concentração externa é baixa. Ao explorar a energia de gradientes iônicos, a célula amplia sua capacidade de acumular substâncias essenciais, integrando diferentes sistemas de transporte em uma rede funcional coordenada.
A manutenção dos gradientes eletroquímicos constitui, portanto, uma das principais funções do transporte ativo. Esses gradientes representam formas de energia potencial que podem ser convertidas em trabalho celular, participando de processos que vão desde a geração de sinais elétricos até a síntese de ATP em organelas como mitocôndrias e cloroplastos. A membrana, nesse contexto, atua como um dispositivo de armazenamento e conversão de energia, reforçando seu papel central na organização celular [Figura 4.18].
Além disso, o transporte ativo contribui para a regulação do volume celular e do equilíbrio osmótico. Ao controlar a entrada e saída de íons, a célula evita o acúmulo excessivo de água ou sua perda, mantendo condições adequadas para a atividade metabólica. Essa função é particularmente importante em organismos submetidos a variações ambientais, nos quais a estabilidade interna depende de mecanismos eficientes de regulação.
A análise do transporte ativo evidencia que a célula não apenas responde passivamente às condições do meio, mas atua de forma ativa na construção e manutenção de sua organização interna. Ao investir energia na geração de gradientes, a célula cria condições para o funcionamento de processos complexos que dependem de diferenças químicas e elétricas. Essa capacidade de operar contra a tendência natural ao equilíbrio é uma das características mais marcantes dos sistemas vivos, preparando o cenário para mecanismos ainda mais sofisticados de transporte e comunicação que serão explorados nas seções seguintes.
4.6 Transporte vesicular: endocitose e exocitose #
A bicamada lipídica, mesmo com a presença de proteínas transportadoras, impõe limitações ao trânsito de partículas de grande porte e de macromoléculas. Para contornar essa restrição, as células desenvolveram mecanismos baseados na deformação e reorganização da própria membrana, permitindo o transporte de materiais por meio de vesículas. Esse processo, conhecido como transporte vesicular, não apenas amplia a capacidade de intercâmbio com o meio, mas também integra a dinâmica interna da célula, conectando diferentes compartimentos membranosos em um sistema contínuo.
A endocitose corresponde ao conjunto de processos pelos quais a célula internaliza substâncias presentes no meio extracelular. Esse mecanismo envolve a invaginação da membrana plasmática, seguida pelo destacamento de uma vesícula que encapsula o material a ser transportado [Figura 4.19]. Dependendo da natureza do material e do mecanismo envolvido, a endocitose pode assumir diferentes formas. A fagocitose caracteriza-se pela ingestão de partículas grandes, como microrganismos ou detritos celulares, sendo típica de células especializadas, como macrófagos. Já a pinocitose envolve a captação de fluido extracelular e de solutos dissolvidos, ocorrendo de maneira mais generalizada em diferentes tipos celulares.
Uma forma particularmente sofisticada de endocitose é a mediada por receptores, na qual moléculas específicas são reconhecidas por proteínas receptoras localizadas na membrana. Após a ligação do ligante ao receptor, regiões da membrana enriquecidas em proteínas de revestimento, como a clatrina, formam invaginações que dão origem a vesículas altamente seletivas [Figura 4.20]. Esse mecanismo permite à célula concentrar e internalizar substâncias específicas, como lipoproteínas e hormônios, mesmo quando presentes em baixas concentrações no meio extracelular.
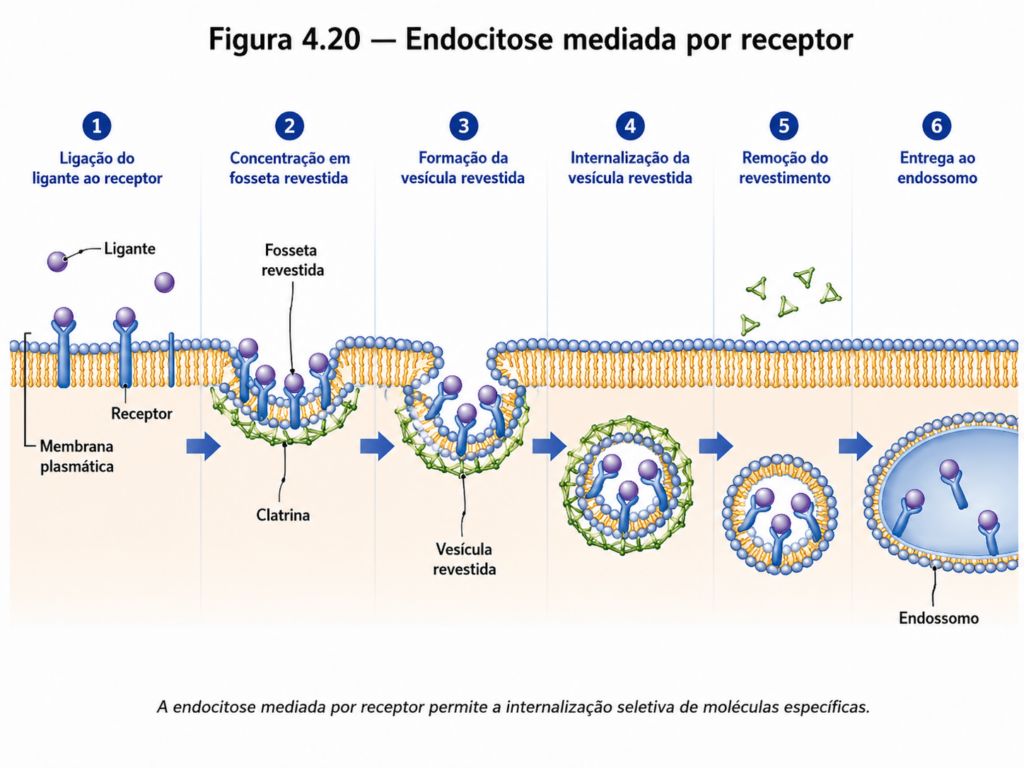
Uma vez internalizadas, as vesículas endocíticas podem seguir diferentes destinos dentro da célula. Frequentemente, elas se fundem com endossomos, compartimentos que atuam na triagem do material internalizado. A partir daí, as moléculas podem ser direcionadas para degradação em lisossomos, recicladas de volta à membrana ou encaminhadas para outros compartimentos celulares [Tabela 4.6]. Esse sistema de triagem evidencia a complexidade do transporte vesicular, que não se limita à internalização, mas envolve decisões funcionais sobre o destino do material transportado.
Em oposição à endocitose, a exocitose corresponde ao processo de liberação de substâncias para o meio extracelular. Nesse caso, vesículas intracelulares, frequentemente originadas do sistema de endomembranas, deslocam-se até a membrana plasmática, com a qual se fundem, liberando seu conteúdo para o exterior [Figura 4.21]. Esse processo é fundamental para a secreção de proteínas, neurotransmissores, hormônios e componentes da matriz extracelular.
A exocitose pode ocorrer de forma constitutiva ou regulada. Na via constitutiva, a secreção ocorre continuamente, sendo responsável pela reposição de componentes da membrana e pela liberação constante de determinadas moléculas. Já na via regulada, a liberação do conteúdo vesicular depende de sinais específicos, como variações na concentração de íons Ca²⁺, permitindo que a célula responda de maneira controlada a estímulos externos [Figura 4.22]. Esse tipo de exocitose é característico de células especializadas, como neurônios e células endócrinas.
A formação, o transporte e a fusão de vesículas envolvem um conjunto altamente coordenado de proteínas que garantem a especificidade e a eficiência do processo. Proteínas de revestimento, como clatrina e caveolina, participam da formação das vesículas, enquanto proteínas de fusão, como as SNAREs, asseguram que vesículas se fundam apenas com os compartimentos corretos. Esse nível de especificidade é essencial para a manutenção da organização celular e para a integração funcional dos sistemas membranosos.
O transporte vesicular também desempenha papel central na renovação da membrana plasmática e na manutenção de sua composição. A constante internalização e reciclagem de componentes permitem ajustes rápidos na quantidade e no tipo de proteínas presentes na superfície celular, influenciando diretamente processos como adesão, sinalização e captação de nutrientes.
A análise da endocitose e da exocitose revela que a membrana plasmática não é uma estrutura estática, mas um sistema em permanente remodelação. A capacidade de formar e fundir vesículas permite à célula expandir significativamente suas possibilidades de interação com o meio e de organização interna. Esse dinamismo reforça o papel da membrana como elemento integrador, conectando processos de transporte, sinalização e metabolismo em uma rede funcional altamente coordenada.
4.7 Especializações da membrana plasmática #
A organização geral da membrana plasmática, baseada na bicamada lipídica e na distribuição dinâmica de proteínas, fornece uma estrutura funcional versátil. No entanto, em células eucariontes, essa organização não é homogênea. A membrana apresenta regiões especializadas que concentram componentes específicos e executam funções diferenciadas, refletindo a necessidade de otimizar processos como absorção, adesão, sinalização e interação com o ambiente. Essas especializações evidenciam que a membrana não atua como uma superfície uniforme, mas como um mosaico funcional altamente organizado.
Um dos níveis mais importantes dessa especialização corresponde à formação de domínios lipídicos, regiões da membrana com composição distinta em relação ao restante da bicamada. Entre esses domínios, destacam-se os chamados “rafts lipídicos”, áreas enriquecidas em colesterol e esfingolipídios que apresentam organização mais ordenada e menor fluidez relativa [Figura 4.23]. Esses microdomínios funcionam como plataformas para a concentração de proteínas envolvidas em processos de sinalização e tráfego membranar, favorecendo a interação entre componentes moleculares que participam de vias específicas.
Além dos domínios lipídicos, a membrana pode apresentar especializações estruturais que aumentam sua superfície ou adaptam sua função a contextos específicos. As microvilosidades são um exemplo clássico desse tipo de modificação. Presentes em células epiteliais absorptivas, como as do intestino, essas projeções aumentam significativamente a área de superfície da membrana, ampliando a capacidade de absorção de nutrientes [Figura 4.24]. Internamente, as microvilosidades são sustentadas por feixes de microfilamentos de actina, evidenciando a integração entre a membrana e o citoesqueleto.
Outro aspecto relevante das especializações de membrana é a polaridade celular, característica marcante de células epiteliais. Nessas células, a membrana plasmática é organizada em domínios distintos — apical, lateral e basal — cada um com composição e função específicas [Figura 4.25]. O domínio apical, voltado para o lúmen ou para o ambiente externo, frequentemente apresenta especializações como microvilosidades, enquanto os domínios laterais e basais estão envolvidos em interações célula-célula e célula-matriz. Essa organização permite a compartimentalização funcional da membrana, essencial para processos como transporte vetorial de substâncias.
As interações da membrana com o citoesqueleto também contribuem para sua especialização funcional. Proteínas de ancoragem conectam componentes da membrana a filamentos de actina, microtúbulos e filamentos intermediários, estabilizando regiões específicas e restringindo a mobilidade de proteínas. Esse ancoramento não apenas confere suporte estrutural, mas também influencia a distribuição de receptores e transportadores, modulando a resposta celular a estímulos [Tabela 4.7].
Além disso, a membrana plasmática estabelece interações com a matriz extracelular e com outras células, formando estruturas especializadas de adesão. Essas regiões são fundamentais para a organização dos tecidos e para a transmissão de sinais mecânicos e químicos. A capacidade de formar e manter essas conexões reforça o papel da membrana como interface ativa entre a célula e seu ambiente.
A existência dessas especializações demonstra que a membrana plasmática é um sistema altamente adaptável, capaz de reorganizar seus componentes de acordo com as demandas funcionais da célula. Essa plasticidade permite a formação de regiões com propriedades distintas dentro de uma mesma membrana, ampliando sua capacidade de executar funções complexas.
A análise das especializações da membrana evidencia, portanto, que sua organização transcende a simples distribuição de lipídios e proteínas. Trata-se de uma arquitetura funcional dinâmica, na qual a localização e a interação dos componentes determinam a eficiência dos processos celulares. Essa organização diferenciada prepara o terreno para a integração funcional da membrana com outros sistemas celulares, consolidando seu papel como elemento central na coordenação das atividades biológicas.
4.8 Integração funcional da membrana com a célula #
A análise isolada da membrana plasmática, ainda que essencial para compreender sua estrutura e propriedades, é insuficiente para captar sua real importância no contexto celular. A membrana não atua de forma independente, mas como um elemento integrador que conecta processos físicos, químicos e biológicos, articulando o interior da célula com o ambiente externo e coordenando interações com outros sistemas intracelulares. Essa integração funcional é o que permite à célula operar como uma unidade organizada, capaz de responder de maneira adaptativa a estímulos internos e externos.
No nível mais imediato, a membrana estabelece a interface entre o meio extracelular e o citoplasma, controlando o fluxo de substâncias e garantindo a manutenção de condições internas compatíveis com a vida. Esse controle não se limita ao transporte, mas envolve também a percepção de sinais provenientes do ambiente. Receptores de membrana reconhecem moléculas específicas e desencadeiam cascatas de sinalização intracelular, permitindo que a célula ajuste seu comportamento em função de alterações externas [Figura 4.26]. Dessa forma, a membrana funciona como um sistema sensorial, capaz de converter informações químicas em respostas biológicas.
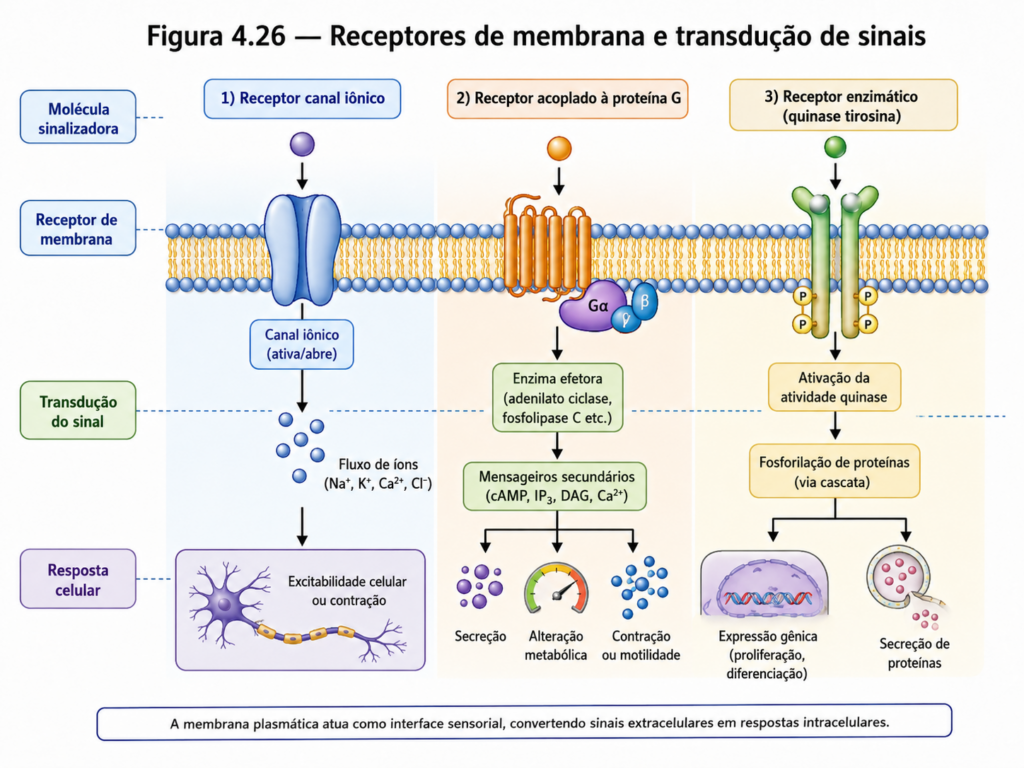
Essa capacidade de percepção e resposta está diretamente relacionada à organização espacial dos componentes da membrana. A distribuição de proteínas e lipídios em domínios específicos permite a formação de complexos funcionais que integram recepção de sinais, transdução e resposta celular. A proximidade entre receptores, proteínas adaptadoras e enzimas facilita a propagação eficiente dos sinais, evidenciando que a arquitetura da membrana é determinante para a eficácia desses processos.
A integração funcional da membrana também se manifesta em sua relação com o citoesqueleto. A ancoragem de proteínas de membrana a filamentos citosqueléticos permite a transmissão de forças mecânicas e a manutenção da forma celular. Além disso, essa conexão regula a mobilidade de proteínas na membrana e contribui para a formação de regiões especializadas. Em células sujeitas a estresse mecânico, como células vegetais ou epiteliais, essa interação é fundamental para a estabilidade estrutural e para a adaptação a variações ambientais [Figura 4.27].
Outro nível de integração ocorre entre a membrana plasmática e o sistema de endomembranas. O transporte vesicular estabelece uma continuidade funcional entre a membrana, o retículo endoplasmático, o complexo de Golgi e outras organelas, permitindo o tráfego direcionado de proteínas e lipídios. Essa rede dinâmica garante que componentes sintetizados no interior da célula sejam corretamente distribuídos e incorporados à membrana ou secretados, mantendo a coerência funcional do sistema celular [Tabela 4.8].
A membrana também participa da organização metabólica da célula, especialmente por meio da manutenção de gradientes eletroquímicos. Esses gradientes não apenas sustentam processos de transporte, mas funcionam como reservas de energia que podem ser utilizadas em diferentes contextos, como na síntese de ATP ou na geração de sinais elétricos. Assim, a membrana atua como um elemento central na gestão energética celular, integrando metabolismo e transporte em um sistema coordenado.
Em organismos multicelulares, a integração funcional da membrana assume uma dimensão adicional, relacionada à comunicação entre células. A presença de receptores, moléculas de adesão e estruturas de junção permite que células interajam entre si, formando tecidos organizados e coordenando respostas coletivas. Essa capacidade de integração intercelular depende diretamente da organização e da funcionalidade da membrana plasmática.
A compreensão da membrana como um elemento integrador revela que sua função vai além da delimitação celular. Ela constitui um sistema dinâmico que articula transporte, sinalização, estrutura e metabolismo, permitindo que a célula opere como uma unidade funcional coerente. Essa visão integrada é fundamental para compreender processos mais complexos que envolvem múltiplos sistemas celulares, consolidando a membrana como um dos principais eixos organizadores da vida celular.
4.9 Perspectivas modernas em biologia de membranas #
A concepção clássica da membrana como uma bicamada lipídica com proteínas inseridas, embora fundamental, tem sido progressivamente refinada à medida que novas abordagens experimentais permitem observar sua organização em escalas espaciais e temporais mais precisas. A membrana, antes descrita como um sistema relativamente homogêneo e fluido, revela-se, em análises contemporâneas, como uma estrutura altamente organizada, dinâmica e heterogênea, na qual interações moleculares complexas determinam funções específicas.
Um dos avanços mais relevantes nesse campo está associado à identificação de nanodomínios funcionais, regiões da membrana cuja organização ocorre em escalas inferiores à resolução da microscopia óptica convencional. Esses nanodomínios, frequentemente associados aos chamados “rafts lipídicos”, apresentam composição distinta, com enriquecimento em colesterol, esfingolipídios e proteínas específicas [Figura 4.28]. Diferentemente de uma distribuição aleatória, a membrana apresenta uma organização hierárquica, na qual essas regiões funcionam como plataformas para a coordenação de processos celulares, especialmente aqueles relacionados à sinalização.
A existência desses domínios sugere que a mobilidade lateral dos componentes da membrana, anteriormente considerada ampla e relativamente livre, é modulada por restrições estruturais e funcionais. Interações com o citoesqueleto, agregação proteica e variações locais na composição lipídica contribuem para a formação de regiões com propriedades distintas. Esse cenário reforça a ideia de que a membrana não é apenas fluida, mas organizada em níveis múltiplos, nos quais a dinâmica molecular é cuidadosamente regulada.
O avanço das técnicas experimentais tem sido decisivo para essa mudança de paradigma. Métodos como recuperação de fluorescência após fotobranqueamento (FRAP) permitiram quantificar a mobilidade de proteínas e lipídios na membrana, enquanto técnicas de microscopia de super-resolução possibilitaram visualizar estruturas em escala nanométrica, revelando padrões de organização anteriormente inacessíveis [Figura 4.29]. Essas abordagens evidenciam que a membrana apresenta uma complexidade estrutural muito maior do que a inicialmente proposta pelos modelos clássicos.
Além disso, a membrana tem sido reconhecida como uma plataforma central para a organização de redes de sinalização celular. A proximidade espacial entre receptores, enzimas e proteínas adaptadoras em regiões específicas da membrana favorece a formação de complexos multiproteicos que amplificam e direcionam sinais intracelulares. Esse arranjo espacial não apenas aumenta a eficiência das respostas celulares, mas também confere especificidade às vias de sinalização, permitindo que diferentes estímulos sejam processados de maneira distinta [Tabela 4.9].
A compreensão moderna das membranas também incorpora sua plasticidade funcional. A composição lipídica e proteica pode ser rapidamente modificada em resposta a estímulos ambientais, permitindo ajustes dinâmicos nas propriedades da membrana. Em organismos vegetais, por exemplo, alterações na proporção de ácidos graxos insaturados modulam a fluidez da membrana em resposta a variações de temperatura, contribuindo para a adaptação a condições adversas [Figura 4.30]. Essa capacidade adaptativa reforça o papel da membrana como elemento central na resposta ao estresse ambiental.
No campo da biotecnologia, o conhecimento aprofundado sobre a organização e a dinâmica das membranas tem gerado aplicações relevantes. A manipulação de lipídios e proteínas de membrana permite o desenvolvimento de sistemas de liberação controlada de fármacos, biossensores e estratégias de engenharia celular. Em sistemas agrícolas, a compreensão da estabilidade de membranas sob estresse hídrico ou térmico tem sido explorada para o desenvolvimento de culturas mais resistentes, evidenciando a importância desse conhecimento para a inovação tecnológica.
A integração desses avanços redefine a compreensão da membrana como uma estrutura multifuncional, na qual organização espacial, dinâmica molecular e interação com outros sistemas celulares convergem para sustentar a funcionalidade biológica. Longe de um modelo estático, a membrana emerge como um sistema adaptativo, capaz de reorganizar-se continuamente em resposta a demandas internas e externas.
Essa perspectiva moderna não substitui os modelos clássicos, mas os amplia, incorporando níveis adicionais de complexidade que refletem a realidade dos sistemas biológicos. A compreensão da membrana, portanto, evolui de uma descrição estrutural para uma visão integrada, na qual sua organização dinâmica é central para a coordenação dos processos celulares, estabelecendo as bases para o entendimento de fenômenos mais complexos que envolvem interação celular, desenvolvimento e adaptação ao ambiente.
Estudo dirigido — Capítulo 4: Membrana Celular #
- Explique como a natureza anfipática dos fosfolipídios permite a formação espontânea da bicamada lipídica em meio aquoso.
- Analise o modelo do mosaico fluido e explique por que ele descreve a membrana plasmática como uma estrutura dinâmica, e não como uma barreira rígida.
- Diferencie proteínas integrais, proteínas periféricas e proteínas ancoradas por lipídios, relacionando cada tipo com possíveis funções na membrana celular.
- Compare difusão simples, difusão facilitada e transporte ativo, destacando o papel do gradiente de concentração, do gradiente eletroquímico e do gasto de energia.
- Explique como a endocitose mediada por receptor permite maior seletividade no transporte de moléculas para o interior da célula.
Pergunta de pesquisa #
- Pesquise como alterações na composição lipídica da membrana plasmática influenciam a tolerância de plantas ao estresse hídrico, salino ou térmico. Relacione sua resposta com fluidez da membrana, integridade celular e adaptação fisiológica das culturas agrícolas.



