Os lipídios desempenham papéis críticos na organização celular e no armazenamento de energia. Este capítulo explora sua natureza hidrofóbica, classificações e funções, com ênfase na formação de membranas biológicas. A bicamada lipídica é apresentada como estrutura dinâmica que regula a interação entre célula e ambiente. Além do papel energético, destaca-se sua participação em processos de sinalização.
5.1 Natureza química dos lipídios e definição funcional #
Ao contrário da maioria das biomoléculas clássicas, cuja identidade química pode ser definida por estruturas relativamente homogêneas e funções bem delimitadas, os lipídios constituem uma classe conceitualmente mais ampla, cuja unidade não reside em uma arquitetura molecular específica, mas sim em um comportamento físico-químico compartilhado. Essa característica fundamental — a baixa solubilidade em água e a alta afinidade por solventes apolares — estabelece o eixo central a partir do qual se compreende a natureza dos lipídios. Em um ambiente dominado pela água, como o interior das células, essa propriedade não representa uma limitação, mas sim um princípio organizador essencial da vida [Figura].
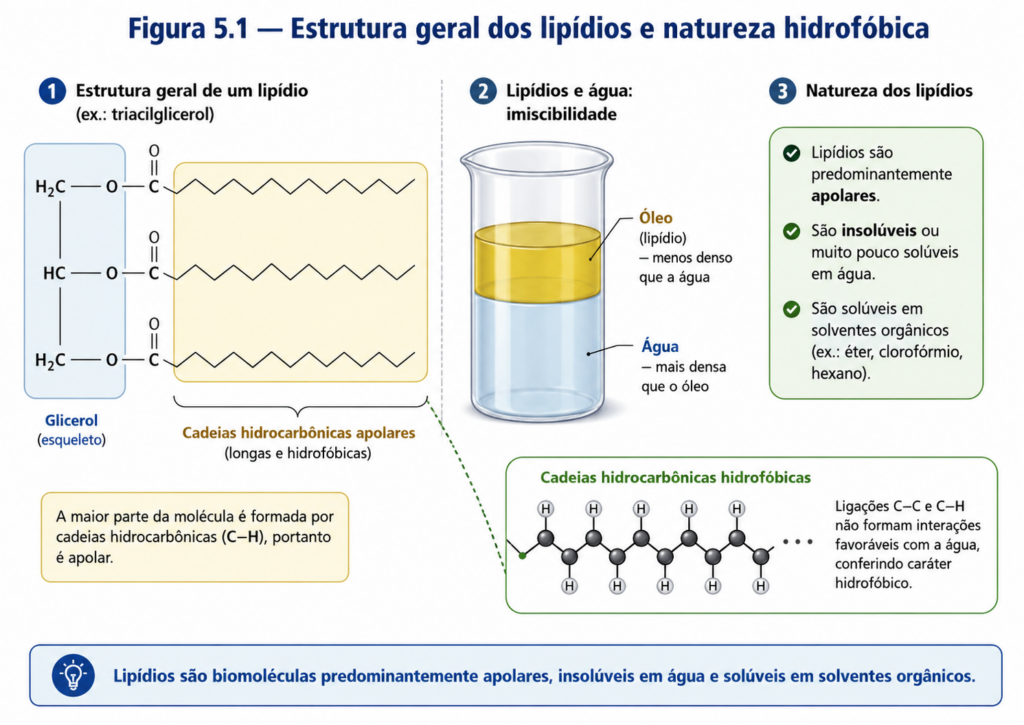
Do ponto de vista químico, os lipídios são compostos predominantemente por cadeias hidrocarbonadas longas ou estruturas ricas em ligações carbono–hidrogênio, o que confere a essas moléculas um caráter apolar. A ausência de grupos funcionais altamente eletronegativos ou ionizáveis reduz sua capacidade de formar interações com a água, especialmente ligações de hidrogênio. Como consequência, quando inseridos em meio aquoso, os lipídios tendem a se organizar de modo a minimizar o contato com a água, fenômeno que está na base do chamado efeito hidrofóbico. Esse efeito, embora não seja uma força no sentido clássico, emerge da reorganização da água ao redor de moléculas apolares e constitui um dos princípios termodinâmicos mais importantes da bioquímica estrutural.
A definição funcional dos lipídios, portanto, transcende a simples descrição estrutural e passa a incorporar seu comportamento em sistemas biológicos. Em vez de serem definidos apenas por “o que são”, os lipídios são mais adequadamente compreendidos por “como se comportam” e “o que fazem” nos contextos celulares. Essa abordagem permite integrar, sob uma mesma categoria, moléculas tão diversas quanto ácidos graxos, triacilgliceróis, fosfolipídios, esfingolipídios e esteroides, que, embora estruturalmente distintos, compartilham propriedades físico-químicas que determinam papéis biológicos convergentes.
Essa diversidade estrutural reflete diretamente a multiplicidade de funções desempenhadas pelos lipídios. Em primeiro lugar, destacam-se como a principal forma de armazenamento de energia em organismos vivos. Devido à alta proporção de ligações carbono–hidrogênio, cuja oxidação libera grande quantidade de energia, os lipídios apresentam densidade energética significativamente superior à dos carboidratos. Essa característica os torna particularmente eficientes como reserva metabólica de longo prazo, permitindo que organismos armazenem grandes quantidades de energia em volumes relativamente pequenos [Tabela].

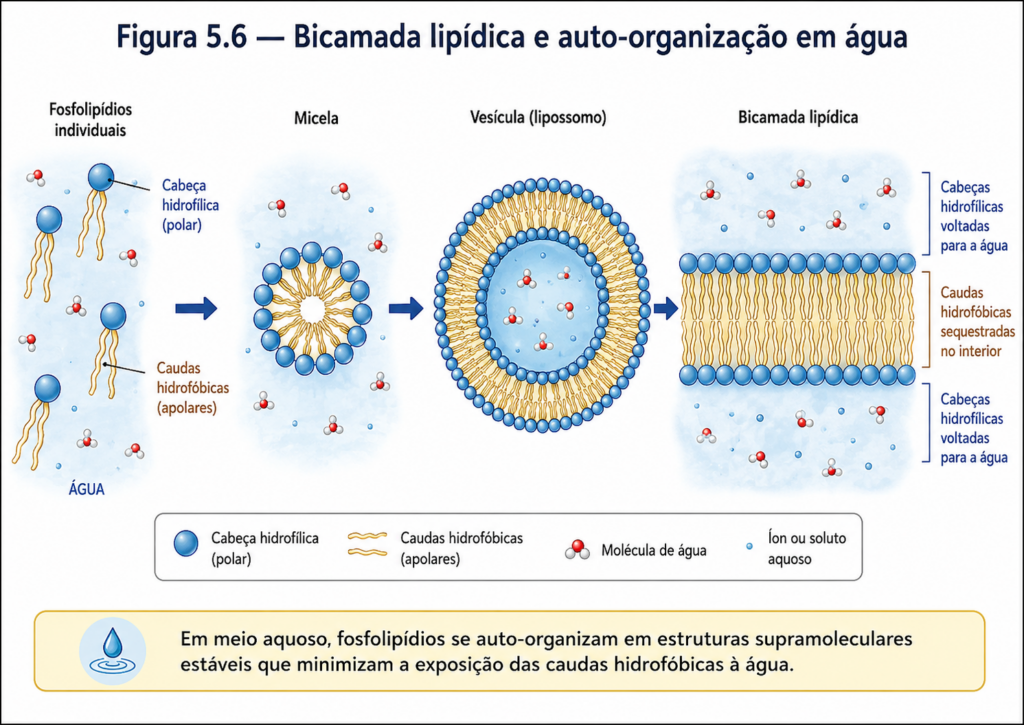
Além do papel energético, os lipídios são elementos estruturais indispensáveis. A formação de membranas biológicas — um dos marcos evolutivos mais importantes na origem da vida — depende diretamente da capacidade de certas classes de lipídios de se organizarem espontaneamente em estruturas supramoleculares, como bicamadas. Nesses arranjos, moléculas anfipáticas, que possuem simultaneamente regiões hidrofílicas e hidrofóbicas, orientam-se de maneira a criar barreiras dinâmicas que delimitam compartimentos celulares, regulam o fluxo de substâncias e permitem a manutenção de gradientes químicos essenciais à vida [Figura].
Outro aspecto fundamental da definição funcional dos lipídios reside em sua participação em processos de sinalização celular. Diversos lipídios atuam como moléculas sinalizadoras ou precursores de mediadores bioativos, participando de cascatas regulatórias que controlam desde respostas inflamatórias até a comunicação intercelular. Nesse contexto, a hidrofobicidade, longe de ser apenas uma propriedade física, torna-se uma vantagem funcional, permitindo que essas moléculas se difundam através de membranas e alcancem alvos intracelulares específicos.
A compreensão dos lipídios, portanto, exige uma mudança de paradigma em relação a outras classes de biomoléculas. Não se trata apenas de reconhecer estruturas químicas isoladas, mas de interpretar como propriedades físico-químicas emergentes — especialmente a hidrofobicidade — moldam a organização da matéria viva. É essa perspectiva que permite compreender por que moléculas aparentemente simples, compostas majoritariamente por carbono e hidrogênio, desempenham papéis tão sofisticados na bioquímica dos sistemas biológicos.
Nesse sentido, os lipídios ocupam uma posição singular na arquitetura molecular da vida. Eles não apenas armazenam energia ou compõem estruturas, mas participam ativamente da construção do próprio ambiente celular no qual todas as demais reações bioquímicas ocorrem. Ao estabelecer interfaces, compartimentos e gradientes, os lipídios criam as condições físicas necessárias para a complexidade biológica emergir. Essa dualidade — entre simplicidade estrutural e profundidade funcional — torna os lipídios uma das classes mais estratégicas para a compreensão integrada da bioquímica.
5.2 Ácidos graxos: estrutura, classificação e propriedades #
Entre as diversas classes de lipídios, os ácidos graxos ocupam uma posição central, não apenas por sua abundância nos sistemas biológicos, mas por representarem a unidade estrutural básica a partir da qual grande parte da diversidade lipídica é construída. Compreender sua organização molecular é, portanto, um passo decisivo para entender como os lipídios armazenam energia, formam membranas e participam de processos regulatórios.
Quimicamente, os ácidos graxos são ácidos carboxílicos de cadeia longa, constituídos por uma extremidade polar — o grupo carboxila (–COOH) — e uma cadeia hidrocarbonada apolar. Essa dualidade confere à molécula um caráter anfipático incipiente: embora a longa cadeia seja predominantemente hidrofóbica, a presença do grupo carboxila permite alguma interação com o meio aquoso, especialmente em pH fisiológico, onde o grupo encontra-se majoritariamente ionizado (–COO⁻) [Figura]. Essa característica é fundamental para a reatividade e para o comportamento desses compostos em sistemas biológicos.
A classificação dos ácidos graxos baseia-se principalmente em dois parâmetros estruturais: o comprimento da cadeia carbônica e o grau de insaturação. Quanto ao comprimento, os ácidos graxos podem variar de cadeias curtas (até 6 carbonos), médias (8 a 12 carbonos) até cadeias longas (14 a 22 carbonos ou mais), sendo estas últimas as mais comuns em organismos eucarióticos. Essa variação não é trivial: o tamanho da cadeia influencia diretamente propriedades como solubilidade, ponto de fusão e destino metabólico.
Entretanto, é no grau de insaturação que reside uma das diferenças mais relevantes do ponto de vista bioquímico. Ácidos graxos saturados possuem apenas ligações simples entre carbonos, o que permite uma conformação linear relativamente rígida. Já os ácidos graxos insaturados apresentam uma ou mais ligações duplas, que introduzem descontinuidades estruturais na cadeia. Em sistemas biológicos, essas ligações duplas ocorrem predominantemente na configuração cis, o que gera uma “curvatura” na molécula, dificultando o empacotamento ordenado entre cadeias adjacentes [Figura].
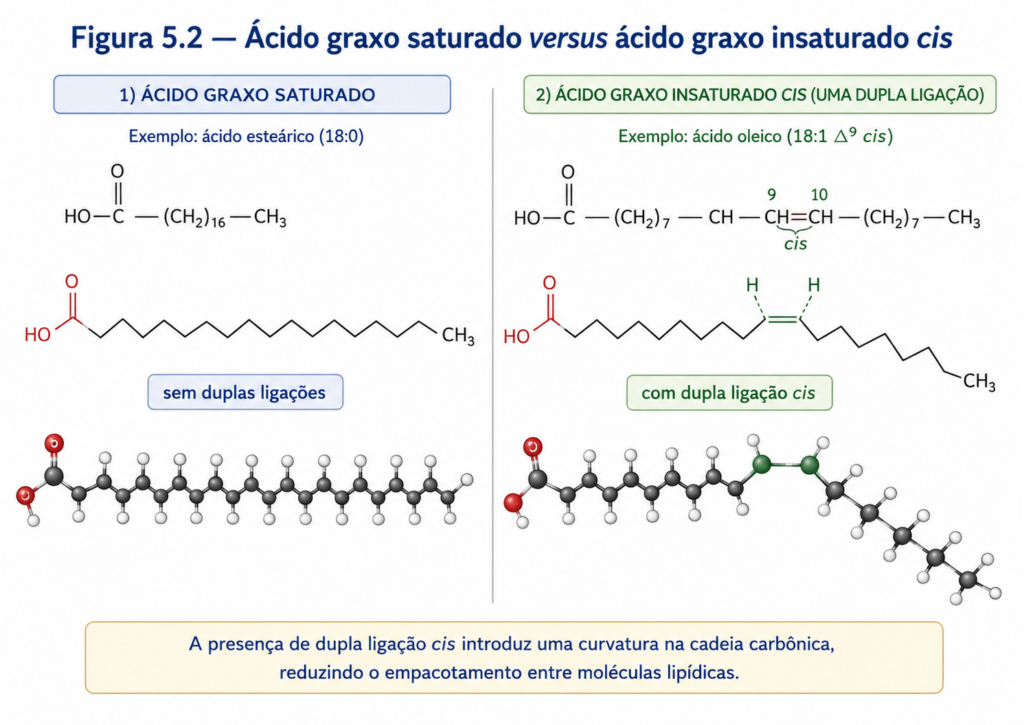
Essa distinção estrutural tem implicações diretas nas propriedades físico-químicas dos lipídios. Cadeias saturadas, por serem mais lineares, interagem de forma mais eficiente por forças de van der Waals, resultando em pontos de fusão mais elevados. Por outro lado, a presença de insaturações — especialmente múltiplas — reduz essas interações, tornando os lipídios mais fluidos em temperaturas fisiológicas. Esse princípio é essencial para a manutenção da fluidez das membranas biológicas, particularmente em organismos expostos a variações térmicas.
Outro aspecto importante refere-se à nomenclatura e à posição das insaturações. Em bioquímica, é comum utilizar o sistema Δ (delta), que indica a posição da dupla ligação a partir da extremidade carboxila, ou o sistema ω (ômega), que a referencia a partir da extremidade metílica. Essa distinção não é meramente descritiva: ácidos graxos da série ω-3 e ω-6, por exemplo, possuem funções fisiológicas específicas e não podem ser sintetizados pelo organismo humano, sendo considerados essenciais e devendo ser obtidos pela dieta.
Do ponto de vista termodinâmico e funcional, a estrutura dos ácidos graxos também determina sua eficiência como moléculas de armazenamento energético. A abundância de ligações carbono–hidrogênio altamente reduzidas faz com que sua oxidação gere grande quantidade de energia livre, o que explica sua utilização como principal reserva energética em muitos organismos. Essa característica, aliada à sua natureza hidrofóbica — que permite armazenamento compacto e sem associação com água — torna os ácidos graxos particularmente eficientes nesse papel.
Além disso, a estrutura dos ácidos graxos influencia diretamente a arquitetura de lipídios mais complexos, como fosfolipídios e triacilgliceróis. Pequenas variações no comprimento da cadeia ou no grau de insaturação podem alterar profundamente propriedades como permeabilidade de membranas, estabilidade estrutural e até mesmo a atividade de proteínas associadas a lipídios. Dessa forma, os ácidos graxos não devem ser vistos apenas como blocos construtivos passivos, mas como elementos determinantes na modulação de sistemas biológicos.
Assim, ao analisar os ácidos graxos sob a perspectiva estrutural e funcional, torna-se evidente que sua importância ultrapassa a simplicidade de sua composição química. Eles representam um elo direto entre a química orgânica básica e a complexidade das estruturas biológicas, sendo fundamentais para a compreensão de como a matéria viva organiza energia, constrói compartimentos e responde às demandas do ambiente.
5.3 Lipídios simples: triacilgliceróis e armazenamento energético #
Se os ácidos graxos representam a unidade estrutural fundamental dos lipídios, é na forma de triacilgliceróis que essa unidade atinge sua máxima eficiência funcional como reserva energética. Essa classe de lipídios simples constitui o principal depósito de energia em organismos vivos, refletindo uma estratégia bioquímica altamente otimizada para armazenamento de carbono reduzido em condições de baixa hidratação.
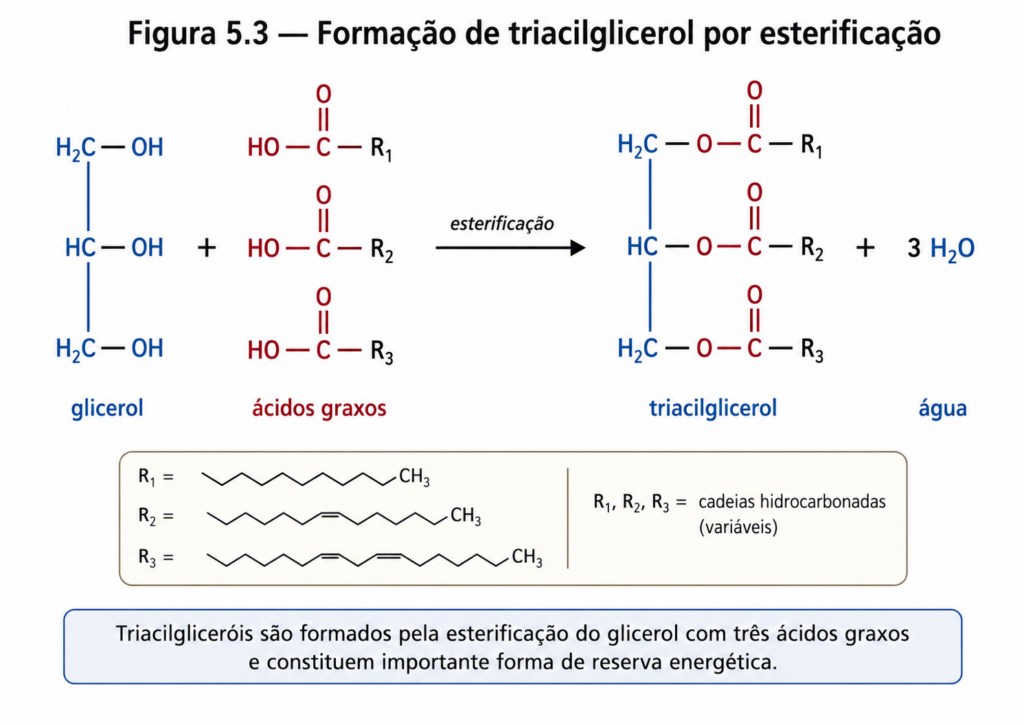
Os triacilgliceróis, também denominados triglicerídeos, são formados pela esterificação de três moléculas de ácidos graxos a uma molécula de glicerol, um álcool de três carbonos. Essa ligação ocorre por meio de reações de condensação, nas quais grupos hidroxila do glicerol reagem com os grupos carboxila dos ácidos graxos, formando ligações éster e liberando moléculas de água [Figura]. O resultado é uma molécula completamente apolar, na qual as propriedades hidrofóbicas das cadeias hidrocarbonadas dominam o comportamento químico.
Essa natureza altamente hidrofóbica tem implicações diretas no modo como os triacilgliceróis são armazenados nos sistemas biológicos. Diferentemente dos carboidratos, que frequentemente se associam à água, os triacilgliceróis são armazenados em forma praticamente anidra, em gotículas lipídicas intracelulares. Essa característica permite um armazenamento extremamente compacto de energia, sem o peso adicional da hidratação. Do ponto de vista físico, trata-se de uma solução eficiente: maior densidade energética por unidade de massa e volume, com menor custo osmótico para a célula.
A superioridade energética dos triacilgliceróis em relação a outras biomoléculas decorre da elevada proporção de átomos de carbono em estado reduzido. Durante a oxidação metabólica, essas cadeias liberam uma quantidade significativamente maior de elétrons, que são transferidos para cofatores redox e, posteriormente, utilizados na produção de ATP. Em termos comparativos, os lipídios fornecem aproximadamente o dobro de energia por grama em relação aos carboidratos, o que explica sua predominância como reserva energética de longo prazo [Tabela].
Além da eficiência energética, os triacilgliceróis desempenham funções adaptativas importantes. Em animais, acumulam-se em tecidos especializados, como o tecido adiposo, onde atuam não apenas como reserva energética, mas também como isolantes térmicos e protetores mecânicos. Em sementes e órgãos de reserva vegetal, os triacilgliceróis garantem o suprimento energético necessário para processos como germinação e crescimento inicial, antes que a fotossíntese esteja plenamente estabelecida.
A composição dos ácidos graxos presentes nos triacilgliceróis também influencia suas propriedades físicas. Misturas ricas em ácidos graxos saturados tendem a formar estruturas mais rígidas, frequentemente sólidas à temperatura ambiente, como observado em gorduras animais. Por outro lado, triacilgliceróis contendo maior proporção de ácidos graxos insaturados apresentam menor ponto de fusão, permanecendo líquidos em condições fisiológicas, como ocorre nos óleos vegetais. Essa variação não é apenas uma curiosidade físico-química, mas reflete adaptações metabólicas e ecológicas específicas.
Do ponto de vista bioquímico, os triacilgliceróis são metabolicamente inertes quando comparados a outras classes de lipídios, como fosfolipídios ou esteroides. Eles não participam diretamente de processos estruturais ou de sinalização celular, sendo mobilizados apenas quando há demanda energética. Essa característica reforça seu papel como forma estável e segura de armazenamento, minimizando interferências com outras funções celulares.
Entretanto, essa mesma eficiência pode se tornar problemática em contextos de desequilíbrio metabólico. O acúmulo excessivo de triacilgliceróis em tecidos não especializados está associado a condições como obesidade e lipotoxicidade, evidenciando que o controle fino do armazenamento e mobilização dessas moléculas é essencial para a homeostase metabólica.
Assim, os triacilgliceróis exemplificam de maneira clara como princípios simples da química — como hidrofobicidade e estado de oxidação do carbono — podem ser explorados pela biologia para resolver problemas complexos, como o armazenamento eficiente de energia. Ao mesmo tempo, revelam que essa eficiência, quando desregulada, pode se tornar um fator crítico na fisiopatologia de diversas doenças, conectando diretamente a bioquímica molecular aos desafios contemporâneos da saúde humana.
5.4 Lipídios complexos: fosfolipídios e esfingolipídios #
Se os triacilgliceróis representam a forma mais eficiente de armazenamento energético, os lipídios complexos emergem como protagonistas na organização estrutural da vida celular. Entre eles, fosfolipídios e esfingolipídios desempenham papel central na construção e funcionalidade das membranas biológicas, estabelecendo a interface dinâmica entre o meio intracelular e o ambiente externo.
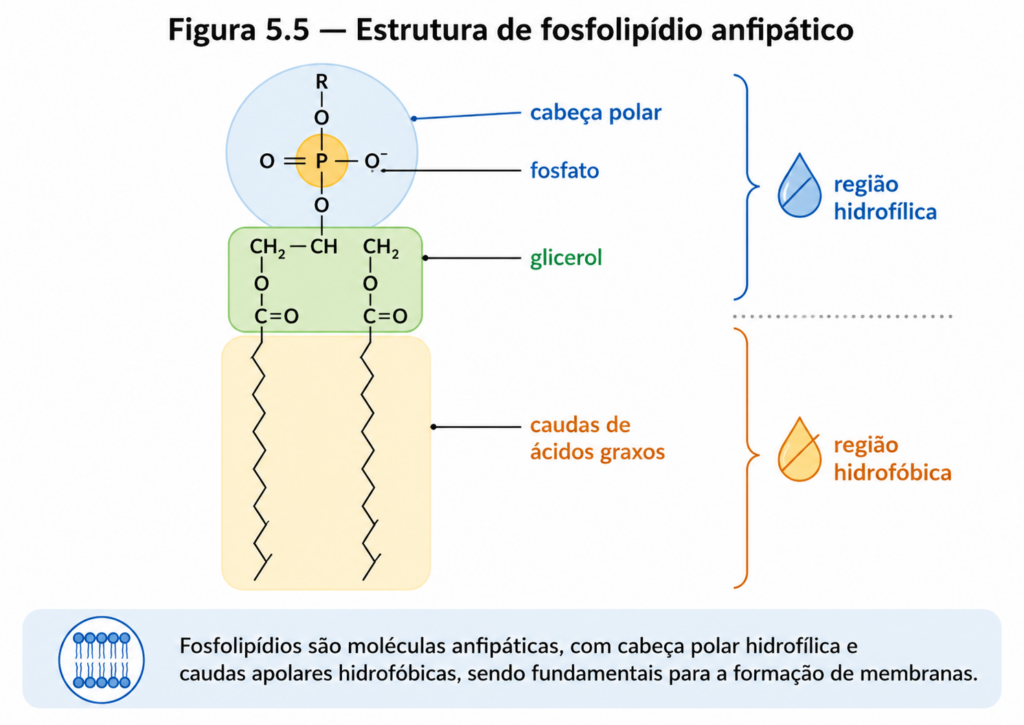
A principal característica que distingue esses lipídios dos lipídios simples é a presença de grupos adicionais ligados à estrutura básica, conferindo-lhes maior complexidade química e, sobretudo, propriedades funcionais ampliadas. Diferentemente dos triacilgliceróis, que são essencialmente apolares, fosfolipídios e esfingolipídios apresentam natureza anfipática bem definida, contendo simultaneamente uma região hidrofóbica — geralmente formada por cadeias de ácidos graxos — e uma região hidrofílica, associada a grupos polares ou carregados [Figura].
Os fosfolipídios constituem a classe mais abundante de lipídios de membrana e são tipicamente derivados do glicerol. Nessa estrutura, duas posições do glicerol estão esterificadas a ácidos graxos, enquanto a terceira está ligada a um grupo fosfato, que por sua vez pode estar associado a diferentes grupos polares, como colina, etanolamina, serina ou inositol. Essa diversidade na cabeça polar gera uma variedade de fosfolipídios com propriedades específicas, capazes de modular características como carga superficial, interação com proteínas e organização da membrana.
A presença do grupo fosfato, fortemente polar e frequentemente ionizado em pH fisiológico, confere aos fosfolipídios uma afinidade significativa por ambientes aquosos. Em contraste, as cadeias hidrocarbonadas permanecem hidrofóbicas, evitando o contato com a água. Essa dualidade estrutural leva à auto-organização espontânea dessas moléculas em sistemas aquosos, resultando na formação de bicamadas lipídicas, micelas ou vesículas, dependendo das condições físico-químicas [Figura]. Esse comportamento não é apenas uma consequência da estrutura molecular, mas um princípio termodinâmico que sustenta a organização celular.
Paralelamente, os esfingolipídios representam uma classe estruturalmente distinta, baseada não no glicerol, mas na esfingosina — um aminoálcool de cadeia longa. A ligação de um ácido graxo à esfingosina forma a ceramida, que serve como núcleo estrutural para diferentes esfingolipídios. A partir dessa base, a adição de grupos polares variados dá origem a subclasses como esfingomielinas e glicoesfingolipídios, estes últimos contendo açúcares em sua estrutura.
Os esfingolipídios não apenas contribuem para a integridade estrutural das membranas, mas também participam ativamente de processos de reconhecimento celular, sinalização e interação célula–célula. Em particular, glicoesfingolipídios presentes na superfície externa das membranas desempenham papéis fundamentais em processos imunológicos e na identificação celular, funcionando como marcadores moleculares específicos.
Do ponto de vista físico, a combinação de fosfolipídios e esfingolipídios nas membranas não é aleatória. Essas moléculas apresentam diferentes graus de saturação, comprimento de cadeia e tamanho de grupos polares, o que influencia diretamente a fluidez, espessura e organização lateral da membrana. Esfingolipídios, por exemplo, tendem a formar regiões mais ordenadas, frequentemente associadas ao colesterol, originando microdomínios funcionais conhecidos como “rafts lipídicos”. Essas regiões são particularmente importantes para a organização de proteínas de sinalização e para a coordenação de respostas celulares.
A complexidade estrutural desses lipídios também se reflete em sua dinâmica metabólica e funcional. Pequenas modificações químicas — como a troca de um grupo polar ou a introdução de uma insaturação — podem alterar significativamente o comportamento da membrana e a interação com proteínas. Isso torna os lipídios complexos elementos altamente reguláveis, capazes de responder a estímulos fisiológicos e ambientais.
Assim, fosfolipídios e esfingolipídios não são meros componentes passivos das membranas, mas agentes ativos na organização e regulação dos sistemas biológicos. Sua capacidade de formar estruturas ordenadas a partir de princípios físico-químicos simples, aliada à diversidade estrutural que possibilita múltiplas funções, evidencia como a bioquímica utiliza variações moleculares sutis para construir sistemas altamente complexos. Nesse contexto, os lipídios complexos representam a transição entre a química das moléculas individuais e a física das estruturas biológicas organizadas, sendo fundamentais para a emergência da vida celular.
5.5 Lipídios de membrana e organização supramolecular #
A emergência da vida celular está diretamente associada à capacidade de determinadas moléculas lipídicas de se organizarem espontaneamente em estruturas ordenadas em meio aquoso. Entre essas estruturas, a bicamada lipídica representa uma das mais importantes inovações da evolução bioquímica, pois estabelece uma barreira seletiva que delimita compartimentos, permite a manutenção de gradientes e cria o ambiente necessário para a ocorrência de reações metabólicas coordenadas. Nesse contexto, os lipídios de membrana deixam de ser apenas entidades químicas individuais e passam a integrar sistemas supramoleculares dinâmicos, nos quais a organização coletiva determina a função [Figura].
A base dessa organização reside na natureza anfipática dos lipídios de membrana, especialmente fosfolipídios e esfingolipídios. Em meio aquoso, essas moléculas não permanecem dispersas de forma aleatória. Ao contrário, tendem a se rearranjar de modo a minimizar a exposição de suas regiões hidrofóbicas à água, enquanto suas cabeças polares interagem com o solvente. Esse comportamento leva à formação espontânea de estruturas como micelas, monocamadas e, sobretudo, bicamadas lipídicas, nas quais duas camadas de lipídios se organizam com as caudas hidrofóbicas voltadas para o interior e as cabeças polares expostas ao meio aquoso em ambos os lados.
Esse processo de auto-organização não requer energia externa direta, sendo conduzido por princípios termodinâmicos associados ao efeito hidrofóbico. A reorganização da água ao redor de superfícies apolares é energeticamente desfavorável; portanto, a agregação das regiões hidrofóbicas reduz essa desordem do sistema, resultando em uma estrutura global mais estável. Assim, a formação da bicamada pode ser entendida como uma consequência inevitável da interação entre moléculas anfipáticas e o ambiente aquoso.
A bicamada lipídica não é uma estrutura rígida, mas sim um sistema altamente dinâmico. Os lipídios que a compõem apresentam mobilidade lateral significativa, podendo difundir-se rapidamente ao longo do plano da membrana. Além disso, rotações e flexões das cadeias hidrocarbonadas ocorrem continuamente, contribuindo para a fluidez da estrutura. Esse caráter dinâmico é essencial para a função biológica das membranas, permitindo reorganizações rápidas em resposta a estímulos e facilitando a interação com proteínas e outras biomoléculas.
Outro aspecto relevante da organização supramolecular das membranas é sua assimetria estrutural. As duas faces da bicamada não são idênticas em composição lipídica, refletindo tanto a origem biossintética dos lipídios quanto processos regulatórios específicos. Essa assimetria tem implicações funcionais importantes, influenciando propriedades como curvatura da membrana, reconhecimento celular e ativação de processos de sinalização.
Além da organização básica em bicamada, as membranas biológicas apresentam níveis adicionais de complexidade. Lipídios com diferentes propriedades físico-químicas podem se distribuir de maneira heterogênea, formando microdomínios com composição distinta. Essas regiões especializadas, frequentemente enriquecidas em esfingolipídios e colesterol, apresentam maior grau de ordenação e desempenham papel crucial na organização de proteínas de membrana e na coordenação de processos de sinalização celular.
A organização supramolecular dos lipídios também está intimamente relacionada à funcionalidade das proteínas de membrana. Muitas dessas proteínas dependem do ambiente lipídico para manter sua estrutura tridimensional e atividade biológica. Alterações na composição lipídica podem modificar a espessura, fluidez ou curvatura da membrana, afetando diretamente a conformação e a função dessas proteínas. Dessa forma, a membrana deve ser entendida como um sistema integrado, no qual lipídios e proteínas interagem de maneira cooperativa.
Do ponto de vista evolutivo, a capacidade de formar membranas estáveis e funcionalmente versáteis foi um passo decisivo na transição de sistemas químicos para sistemas biológicos organizados. A compartimentalização permitiu a especialização de funções, a concentração de reagentes e o estabelecimento de gradientes eletroquímicos, elementos fundamentais para a complexidade metabólica observada nos organismos atuais.
Assim, a organização supramolecular dos lipídios de membrana representa muito mais do que um arranjo estrutural: trata-se de um princípio organizador da vida. A partir de interações físico-químicas simples, emergem estruturas capazes de sustentar processos altamente complexos, evidenciando como a bioquímica transforma propriedades moleculares em sistemas funcionais integrados.
5.6 Fluidez de membrana e fatores moduladores #
A funcionalidade das membranas biológicas não depende apenas de sua existência como barreiras estruturais, mas sobretudo de sua capacidade de manter um estado físico adequado — nem rígido demais, nem excessivamente fluido. Esse equilíbrio é descrito pelo conceito de fluidez de membrana, uma propriedade emergente que resulta da organização e das interações entre os lipídios que compõem a bicamada. A fluidez determina diretamente processos essenciais como difusão lateral de moléculas, atividade de proteínas de membrana, transporte de solutos e resposta a estímulos ambientais [Figura].
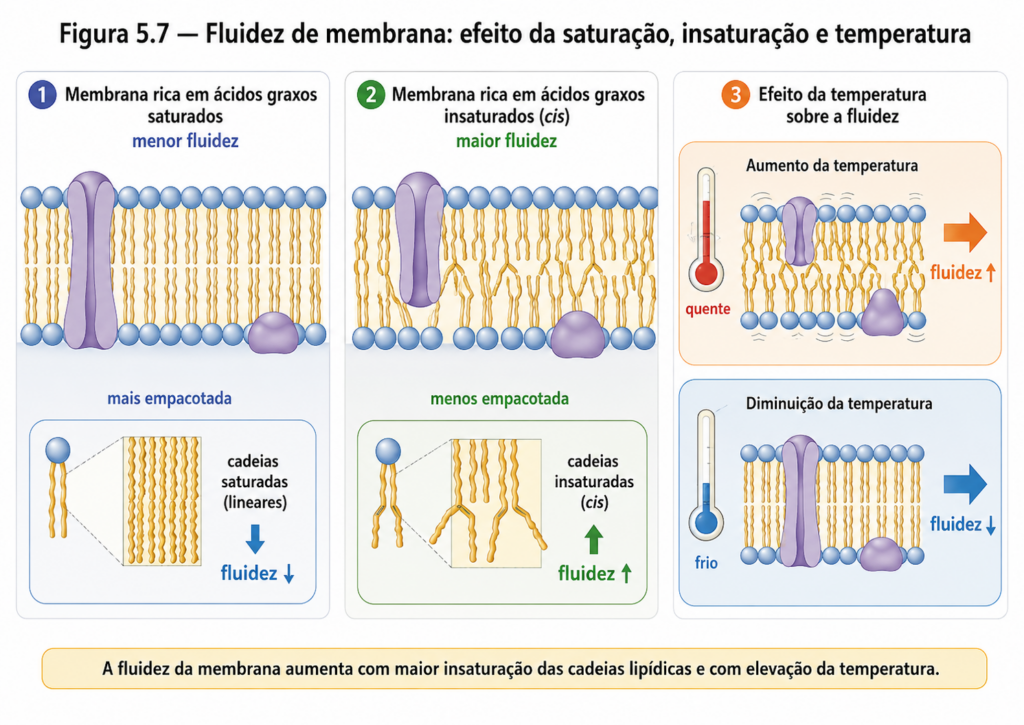
Do ponto de vista físico, a fluidez pode ser entendida como o grau de mobilidade das moléculas lipídicas dentro da bicamada. Em condições nas quais as interações entre cadeias hidrocarbonadas são intensas e bem organizadas, a membrana tende a um estado mais ordenado, frequentemente descrito como “fase gel”. Nesse estado, o movimento molecular é restrito, comprometendo a dinâmica da membrana. Em contraste, quando essas interações são reduzidas, a bicamada assume um estado mais desordenado e fluido, denominado “fase líquida cristalina”, no qual os lipídios apresentam maior liberdade de movimento.
Entre os fatores que modulam essa propriedade, a temperatura desempenha papel fundamental. À medida que a temperatura aumenta, a energia cinética das moléculas também se eleva, reduzindo as interações intermoleculares e promovendo maior fluidez. Em temperaturas mais baixas, ocorre o inverso: as cadeias lipídicas tornam-se mais organizadas, favorecendo a rigidez. Cada membrana apresenta, portanto, uma temperatura de transição característica, na qual ocorre a mudança entre estados mais ordenados e mais fluidos [Tabela].
Entretanto, a composição lipídica da membrana é o principal determinante da sua fluidez em condições fisiológicas. O grau de saturação dos ácidos graxos é particularmente relevante. Cadeias saturadas, por sua conformação linear, permitem um empacotamento mais eficiente, aumentando as interações de van der Waals e promovendo maior rigidez. Por outro lado, cadeias insaturadas, especialmente na configuração cis, introduzem “dobras” na estrutura, dificultando o empacotamento e aumentando a fluidez da membrana. Assim, a proporção entre lipídios saturados e insaturados constitui um mecanismo crucial de regulação.
O comprimento das cadeias hidrocarbonadas também influencia essa propriedade. Cadeias mais longas apresentam maior área de contato intermolecular, intensificando as forças de atração e favorecendo estados mais rígidos. Em contraste, cadeias mais curtas reduzem essas interações, promovendo maior mobilidade. Dessa forma, pequenas variações estruturais nos ácidos graxos podem resultar em mudanças significativas no comportamento físico da membrana.
Um dos moduladores mais importantes da fluidez em células eucarióticas é o colesterol. Sua estrutura rígida, composta por anéis esteroides, permite que ele se insira entre as cadeias de ácidos graxos dos fosfolipídios, exercendo um efeito dual. Em temperaturas elevadas, o colesterol restringe o movimento das cadeias lipídicas, reduzindo a fluidez excessiva. Em temperaturas mais baixas, ele impede o empacotamento ordenado das cadeias, evitando a transição para estados rígidos. Assim, o colesterol atua como um “amortecedor” físico, estabilizando a fluidez da membrana em uma faixa funcional adequada [Figura].
Essa capacidade de ajuste fino da fluidez é particularmente importante em organismos expostos a variações ambientais. Bactérias e plantas, por exemplo, podem alterar a composição de seus lipídios de membrana em resposta a mudanças de temperatura, aumentando o grau de insaturação em ambientes frios para manter a fluidez. Esse fenômeno, conhecido como adaptação homeoviscosa, ilustra como a bioquímica integra propriedades físico-químicas com necessidades fisiológicas.
A fluidez da membrana também tem impacto direto na atividade de proteínas associadas. Enzimas de membrana, canais iônicos e receptores dependem de um ambiente lipídico adequado para manter sua conformação e funcionalidade. Alterações na fluidez podem modificar a dinâmica dessas proteínas, afetando processos como transporte, sinalização e metabolismo. Nesse sentido, a membrana não deve ser vista como um suporte passivo, mas como um componente ativo na regulação da atividade celular.
Do ponto de vista fisiopatológico, alterações na fluidez de membrana estão associadas a diversas condições. Mudanças na composição lipídica podem comprometer a integridade da membrana, afetar a comunicação celular e contribuir para o desenvolvimento de doenças metabólicas e degenerativas. Assim, a manutenção da fluidez adequada não é apenas uma questão estrutural, mas um requisito essencial para a homeostase celular.
Em síntese, a fluidez de membrana emerge como uma propriedade crítica que conecta a estrutura molecular dos lipídios à funcionalidade dos sistemas biológicos. Através da modulação de fatores como temperatura, grau de insaturação, comprimento das cadeias e presença de colesterol, os organismos são capazes de ajustar dinamicamente suas membranas, garantindo a estabilidade e a eficiência dos processos vitais.
5.7 Colesterol e esteroides: estrutura e funções #
Entre os lipídios de membrana, poucos exercem influência tão ampla e multifacetada quanto o colesterol. Diferentemente dos fosfolipídios e esfingolipídios, cuja estrutura é dominada por cadeias hidrocarbonadas flexíveis, o colesterol apresenta um núcleo rígido formado por quatro anéis fusionados — a estrutura esteroidal — que confere propriedades físico-químicas singulares. Essa arquitetura molecular, composta por um pequeno grupo hidroxila polar e uma extensa porção hidrofóbica, posiciona o colesterol como uma molécula anfipática atípica, capaz de interagir simultaneamente com regiões polares e apolares da membrana [Figura].
Do ponto de vista estrutural, o colesterol se organiza na bicamada lipídica de maneira altamente específica. O grupo hidroxila orienta-se próximo às cabeças polares dos fosfolipídios, interagindo com o meio aquoso, enquanto o núcleo esteroidal e a curta cauda hidrocarbonada se inserem entre as cadeias de ácidos graxos. Essa disposição permite que o colesterol module diretamente o arranjo das moléculas vizinhas, influenciando propriedades fundamentais da membrana.
A principal função do colesterol nas membranas biológicas está relacionada à regulação da fluidez. Como discutido anteriormente, a fluidez depende do equilíbrio entre mobilidade e organização das cadeias lipídicas. O colesterol atua como um modulador bidirecional: em temperaturas elevadas, sua estrutura rígida restringe o movimento das cadeias de ácidos graxos, reduzindo a fluidez excessiva; em temperaturas mais baixas, impede o empacotamento ordenado dessas cadeias, evitando a rigidez extrema. Esse comportamento confere às membranas maior estabilidade física, mantendo suas propriedades dentro de uma faixa funcional adequada [Figura].
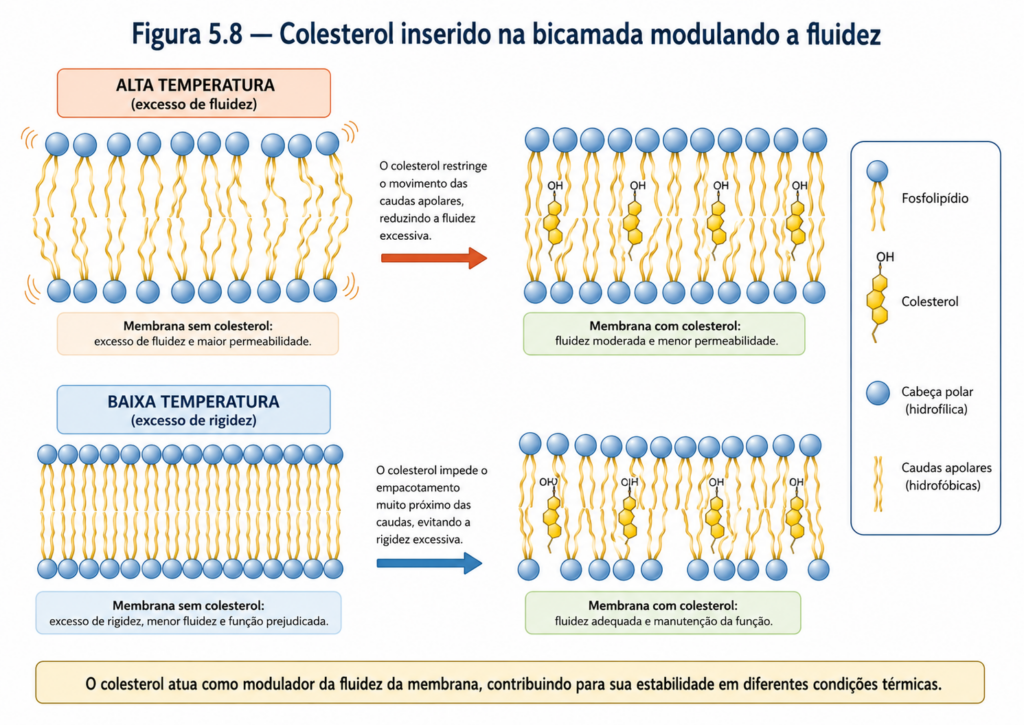
Além da fluidez, o colesterol também contribui para a redução da permeabilidade da membrana a pequenas moléculas polares. Ao preencher espaços entre os fosfolipídios, ele diminui a formação de lacunas transitórias na bicamada, dificultando a passagem não controlada de substâncias. Essa função é particularmente importante em células eucarióticas, nas quais o controle rigoroso do ambiente intracelular é essencial para a manutenção da homeostase.
Outro aspecto relevante é a participação do colesterol na formação de microdomínios de membrana, frequentemente associados a esfingolipídios. Esses domínios, conhecidos como “rafts lipídicos”, apresentam maior grau de ordenação e servem como plataformas para a organização de proteínas envolvidas em processos de sinalização celular. Nesses contextos, o colesterol não atua apenas como modulador físico, mas como elemento organizador de complexos funcionais.
Para além de seu papel estrutural, o colesterol é também um precursor biossintético de uma série de moléculas biologicamente ativas. Entre elas, destacam-se os hormônios esteroides, como os glicocorticoides, mineralocorticoides e hormônios sexuais, que regulam processos fisiológicos fundamentais, incluindo metabolismo, equilíbrio hidroeletrolítico e reprodução. A transformação do colesterol nesses compostos envolve modificações enzimáticas específicas que alteram sua estrutura, conferindo novas propriedades funcionais.
Adicionalmente, o colesterol dá origem aos ácidos biliares, moléculas essenciais para a digestão e absorção de lipídios no trato gastrointestinal. Esses compostos atuam como agentes emulsificantes, facilitando a ação de enzimas lipolíticas e promovendo a solubilização de lipídios em meio aquoso. Assim, o colesterol conecta diretamente a bioquímica estrutural das membranas com processos digestivos e metabólicos sistêmicos.
Do ponto de vista fisiológico, a homeostase do colesterol é rigidamente controlada, envolvendo equilíbrio entre síntese endógena, ingestão dietética, transporte e excreção. Lipoproteínas plasmáticas desempenham papel central nesse processo, transportando colesterol entre tecidos e órgãos. Alterações nesse sistema podem levar ao acúmulo de colesterol em locais inadequados, contribuindo para o desenvolvimento de doenças cardiovasculares, como a aterosclerose.
Essa dualidade — essencialidade biológica e potencial patológico — torna o colesterol uma molécula de grande relevância clínica. Em níveis adequados, ele é indispensável para a integridade estrutural das membranas e para a síntese de moléculas reguladoras. Em excesso ou distribuído de forma inadequada, pode se tornar um fator de risco significativo para doenças crônicas.
Assim, o estudo do colesterol e dos esteroides revela de maneira exemplar como uma única estrutura molecular pode desempenhar múltiplas funções, integrando diferentes níveis de organização biológica. A partir de um núcleo químico relativamente simples, a bioquímica constrói uma rede de processos que conectam a arquitetura das membranas, a regulação hormonal e a fisiologia sistêmica, evidenciando a complexidade funcional que emerge da estrutura molecular.
5.8 Lipídios como moléculas de sinalização #
A compreensão tradicional dos lipídios como meros componentes estruturais ou reservas energéticas revela-se incompleta diante de sua participação ativa em processos de comunicação celular. Em sistemas biológicos complexos, os lipídios assumem funções dinâmicas como mediadores de sinalização, integrando estímulos externos e internos em respostas bioquímicas coordenadas. Essa atuação depende diretamente de suas propriedades físico-químicas, especialmente sua hidrofobicidade e sua capacidade de interação com membranas e proteínas específicas [Figura].
Diferentemente de mensageiros hidrossolúveis, muitos lipídios sinalizadores atuam no interior ou na interface das membranas, onde são gerados e exercem seus efeitos localmente. Essa característica confere rapidez e especificidade à transmissão do sinal, uma vez que a difusão lateral na bicamada permite o alcance eficiente de proteínas-alvo. Entre os principais grupos de lipídios envolvidos nesses processos destacam-se os eicosanoides, os fosfoinositídeos e diversos derivados lipídicos que atuam como segundos mensageiros.
Os eicosanoides constituem uma classe importante de mediadores lipídicos derivados de ácidos graxos poli-insaturados, especialmente o ácido araquidônico. A partir desse precursor, diferentes vias enzimáticas dão origem a moléculas como prostaglandinas, tromboxanos e leucotrienos, cada uma com funções fisiológicas específicas. Esses compostos atuam principalmente em processos inflamatórios, regulação da pressão arterial, agregação plaquetária e resposta imune. Sua ação é tipicamente local e de curta duração, o que permite um controle fino e transitório das respostas celulares [Figura].
Outro sistema fundamental envolve os fosfoinositídeos, lipídios de membrana que, embora presentes em pequenas quantidades, desempenham papéis centrais na sinalização intracelular. A hidrólise de fosfatidilinositol 4,5-bifosfato (PIP₂), por exemplo, gera dois importantes segundos mensageiros: o inositol trifosfato (IP₃), que promove a liberação de cálcio a partir de compartimentos intracelulares, e o diacilglicerol (DAG), que permanece na membrana e ativa proteínas quinases específicas. Esse mecanismo ilustra como a modificação de um lipídio estrutural pode desencadear uma cascata de eventos celulares amplamente amplificada.
Além desses sistemas clássicos, diversos lipídios bioativos atuam diretamente na regulação de processos celulares por interação com receptores específicos. Ácidos graxos livres, esfingolipídios e seus derivados, como ceramidas e esfingosina-1-fosfato, participam da regulação de processos como proliferação celular, apoptose e diferenciação. Essas moléculas funcionam como verdadeiros moduladores metabólicos, conectando o estado bioquímico da célula às suas respostas fisiológicas.
A hidrofobicidade dos lipídios também permite que alguns deles atravessem membranas celulares e atuem como sinalizadores intracelulares de longo alcance. Hormônios esteroides, derivados do colesterol, exemplificam esse comportamento. Por serem solúveis em lipídios, difundem-se facilmente através da bicamada e interagem com receptores intracelulares, modulando diretamente a expressão gênica. Esse mecanismo contrasta com sistemas de sinalização baseados em receptores de membrana, evidenciando a diversidade funcional dos lipídios nesse contexto.
Outro aspecto relevante é a organização espacial da sinalização lipídica. A formação de microdomínios de membrana, enriquecidos em determinados lipídios, cria plataformas especializadas onde proteínas sinalizadoras podem ser recrutadas e ativadas. Esses domínios funcionam como centros de integração de sinais, aumentando a eficiência e a especificidade das respostas celulares.
Do ponto de vista regulatório, a sinalização mediada por lipídios é altamente controlada, envolvendo síntese rápida, modificação enzimática e degradação eficiente dos mediadores. Essa dinâmica garante que os sinais sejam transitórios e precisamente modulados, evitando respostas excessivas ou prolongadas que poderiam comprometer a homeostase celular.
A relevância desses processos torna-se ainda mais evidente em contextos fisiopatológicos. Alterações na produção ou na ação de lipídios sinalizadores estão associadas a diversas doenças, incluindo inflamações crônicas, distúrbios cardiovasculares, câncer e doenças neurodegenerativas. Assim, esses compostos não apenas desempenham papéis essenciais na fisiologia normal, mas também representam alvos importantes para intervenções terapêuticas.
Em síntese, os lipídios como moléculas de sinalização exemplificam a capacidade da bioquímica de transformar propriedades estruturais em funções regulatórias sofisticadas. A partir de moléculas inicialmente associadas à estrutura e ao armazenamento, emergem sistemas complexos de comunicação celular, capazes de integrar múltiplos estímulos e coordenar respostas biológicas em diferentes níveis de organização.
5.9 Digestão, absorção e transporte de lipídios (visão geral) #
A natureza hidrofóbica dos lipídios, que lhes confere vantagens estruturais e energéticas no contexto celular, impõe um desafio fundamental no ambiente fisiológico: como moléculas insolúveis em água podem ser eficientemente digeridas, absorvidas e transportadas em um sistema predominantemente aquoso? A resposta a essa questão envolve uma sequência coordenada de processos físico-químicos e enzimáticos que transformam agregados lipídicos em formas assimiláveis, permitindo sua integração ao metabolismo sistêmico [Figura].
A digestão dos lipídios inicia-se de maneira limitada ainda na cavidade oral e no estômago, mas ocorre predominantemente no intestino delgado, onde condições adequadas de pH e a presença de emulsificantes naturais tornam o processo altamente eficiente. O primeiro passo crítico não é enzimático, mas físico: a emulsificação. Grandes massas lipídicas, inicialmente organizadas em gotículas, são fragmentadas em partículas menores pela ação mecânica e, sobretudo, pela ação dos sais biliares, moléculas anfipáticas derivadas do colesterol. Esses compostos reduzem a tensão superficial entre a fase lipídica e o meio aquoso, estabilizando pequenas gotículas e aumentando significativamente a área de contato para ação enzimática.
Com a emulsificação estabelecida, enzimas lipolíticas passam a atuar de forma mais eficiente. A principal delas, a lipase pancreática, catalisa a hidrólise das ligações éster dos triacilgliceróis, liberando ácidos graxos livres e monoacilgliceróis [Figura]. Esse processo é auxiliado por proteínas acessórias, como a colipase, que estabiliza a interação da lipase com a superfície das gotículas lipídicas emulsificadas. Paralelamente, outras enzimas atuam sobre diferentes classes de lipídios, como fosfolipases e esterases de colesterol, ampliando a gama de produtos gerados.
Os produtos da digestão lipídica não permanecem livres no meio intestinal. Devido à sua baixa solubilidade, são incorporados a estruturas conhecidas como micelas mistas, formadas pela associação com sais biliares. Nessas estruturas, as regiões hidrofóbicas dos lipídios permanecem protegidas no interior, enquanto as porções polares interagem com o meio aquoso, permitindo a difusão dessas partículas até a superfície das células epiteliais intestinais [Figura]. Esse arranjo é essencial para superar a barreira aquosa que recobre o epitélio intestinal.
A absorção propriamente dita ocorre quando os componentes lipídicos deixam as micelas e atravessam a membrana plasmática dos enterócitos. Esse processo pode envolver difusão passiva, facilitada pela natureza lipofílica das moléculas, bem como mecanismos mediados por transportadores específicos. Uma vez no interior da célula, os ácidos graxos e monoacilgliceróis são rapidamente reesterificados, formando novamente triacilgliceróis. Esse passo é crucial para manter o gradiente de absorção e evitar o acúmulo de formas livres potencialmente detergentes.
Os lipídios reesterificados são então incorporados em complexos macromoleculares denominados quilomícrons, que incluem, além de triacilgliceróis, colesterol, fosfolipídios e proteínas específicas chamadas apolipoproteínas. Esses complexos permitem o transporte dos lipídios em meio aquoso, resolvendo o problema da insolubilidade. Devido ao seu tamanho, os quilomícrons não entram diretamente na circulação sanguínea, sendo inicialmente liberados no sistema linfático e, posteriormente, alcançando a corrente sanguínea sistêmica [Figura].
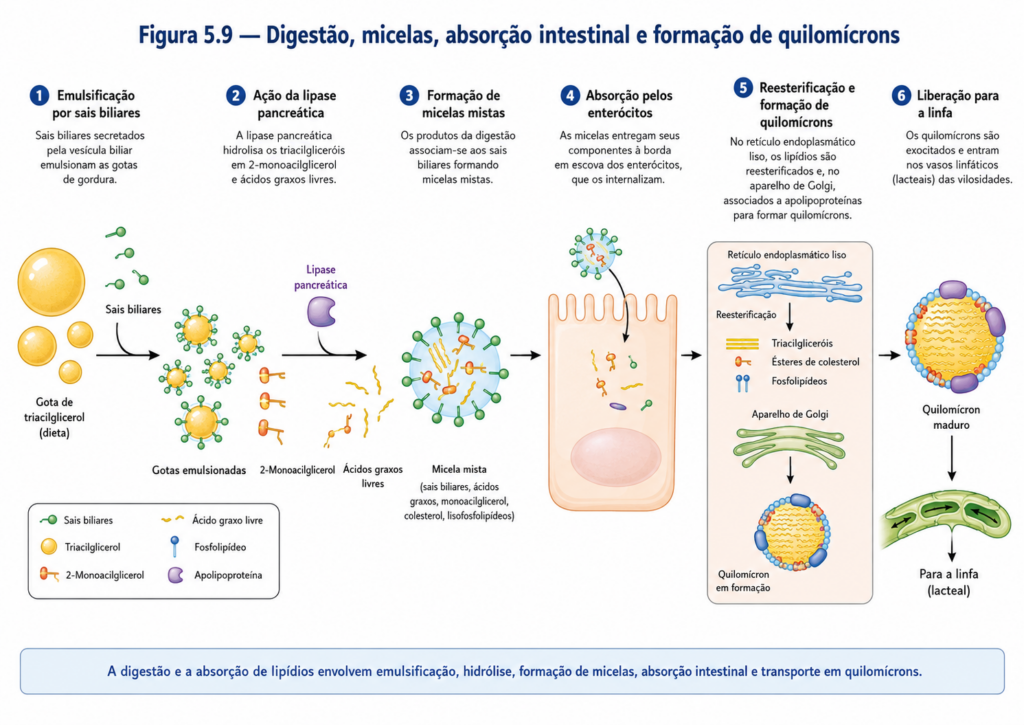
Esse mecanismo de transporte evidencia um princípio central da bioquímica dos lipídios: a necessidade de estruturas intermediárias que permitam sua mobilização em ambientes aquosos. Ao longo do organismo, diferentes tipos de lipoproteínas assumem essa função, distribuindo lipídios entre tecidos conforme as demandas metabólicas.
Do ponto de vista integrativo, a digestão e absorção de lipídios não representam apenas processos isolados, mas etapas iniciais de uma rede metabólica mais ampla. A eficiência desses processos influencia diretamente o estado energético do organismo, a composição das membranas celulares e a disponibilidade de moléculas sinalizadoras. Alterações em qualquer etapa — desde a produção de sais biliares até a formação de lipoproteínas — podem comprometer a homeostase lipídica, resultando em distúrbios nutricionais e metabólicos.
Assim, o processamento dos lipídios no sistema digestório exemplifica de maneira clara como a bioquímica resolve desafios impostos pela própria natureza das moléculas. Através da integração de princípios físicos, químicos e biológicos, o organismo transforma substâncias hidrofóbicas em componentes plenamente funcionais dentro de um ambiente aquoso, garantindo sua utilização eficiente em diferentes contextos fisiológicos.
5.10 Lipoproteínas e transporte sistêmico #
Uma vez absorvidos no intestino e reorganizados em formas compatíveis com o meio aquoso, os lipídios precisam ser distribuídos de maneira eficiente entre os diferentes tecidos do organismo. Esse processo não é trivial: a hidrofobicidade que caracteriza os lipídios impede sua circulação livre no plasma. A solução bioquímica para esse problema é a formação de lipoproteínas — complexos supramoleculares altamente organizados que permitem o transporte sistêmico de lipídios em um ambiente essencialmente aquoso [Figura].
As lipoproteínas são partículas esféricas compostas por um núcleo hidrofóbico, formado principalmente por triacilgliceróis e ésteres de colesterol, envolto por uma monocamada de fosfolipídios, colesterol livre e proteínas específicas denominadas apolipoproteínas. Essa organização é funcionalmente elegante: as regiões hidrofóbicas permanecem protegidas no interior da partícula, enquanto as superfícies polares interagem com o plasma, garantindo estabilidade e solubilidade.
As apolipoproteínas desempenham papéis críticos na dinâmica dessas partículas. Além de contribuírem para a estabilidade estrutural, atuam como sinais de reconhecimento para receptores celulares e como cofatores para enzimas envolvidas no metabolismo lipídico. Assim, as lipoproteínas não são meros veículos passivos, mas sistemas regulados que integram transporte, reconhecimento e transformação metabólica.
Do ponto de vista funcional, as lipoproteínas são classificadas com base em sua densidade, que reflete a proporção relativa entre lipídios e proteínas. Entre as principais classes, destacam-se os quilomícrons, as lipoproteínas de muito baixa densidade (VLDL), de baixa densidade (LDL) e de alta densidade (HDL). Cada uma dessas classes possui composição, origem e função específicas no transporte lipídico sistêmico.
Os quilomícrons, formados nos enterócitos após a absorção intestinal, são responsáveis pelo transporte inicial de lipídios da dieta, especialmente triacilgliceróis, para os tecidos periféricos. Ao circularem pela corrente sanguínea, esses complexos interagem com a lipoproteína lipase, uma enzima localizada na superfície de células endoteliais, que hidrolisa os triacilgliceróis, liberando ácidos graxos para captação pelos tecidos, como músculo e tecido adiposo. À medida que perdem conteúdo lipídico, os quilomícrons são convertidos em remanescentes, que são posteriormente captados pelo fígado.
O fígado, por sua vez, desempenha papel central na redistribuição de lipídios endógenos, principalmente por meio da secreção de VLDL. Essas partículas transportam triacilgliceróis sintetizados no fígado para tecidos periféricos. De maneira análoga aos quilomícrons, as VLDL sofrem ação da lipoproteína lipase, reduzindo seu conteúdo lipídico e dando origem a partículas de densidade intermediária e, posteriormente, às LDL.
As LDL são particularmente relevantes do ponto de vista fisiológico e clínico, pois atuam como principais transportadoras de colesterol para os tecidos. A internalização das LDL pelas células ocorre por meio de receptores específicos que reconhecem apolipoproteínas presentes na superfície dessas partículas. Esse mecanismo permite que o colesterol seja utilizado na síntese de membranas, hormônios esteroides e outras moléculas essenciais. No entanto, quando esse sistema é desregulado, o acúmulo de LDL na circulação pode levar à deposição de colesterol nas paredes arteriais, contribuindo para o desenvolvimento de aterosclerose.
Em contraste, as HDL exercem função protetora, participando do chamado transporte reverso de colesterol. Essas partículas captam o excesso de colesterol dos tecidos periféricos e o transportam de volta ao fígado, onde pode ser metabolizado ou excretado. Esse processo é fundamental para a manutenção do equilíbrio lipídico e para a prevenção do acúmulo patológico de colesterol.
A dinâmica entre essas diferentes classes de lipoproteínas evidencia um sistema altamente integrado, no qual o transporte de lipídios não é linear, mas sim um fluxo contínuo de redistribuição, modificação e regulação. Alterações em qualquer componente desse sistema — seja na síntese, na composição das apolipoproteínas ou na atividade enzimática — podem resultar em distúrbios metabólicos significativos, como dislipidemias e doenças cardiovasculares.
Do ponto de vista bioquímico, as lipoproteínas representam uma solução sofisticada para um problema fundamental: como transportar moléculas hidrofóbicas em um meio aquoso sem comprometer sua funcionalidade. Mais do que isso, constituem um sistema regulatório que conecta a absorção intestinal, o metabolismo hepático e as demandas energéticas e estruturais dos tecidos periféricos.
Assim, o estudo das lipoproteínas revela não apenas os mecanismos de transporte lipídico, mas também a complexa rede de interações que sustenta a homeostase metabólica. A partir da organização molecular dessas partículas, emerge um sistema dinâmico capaz de integrar diferentes níveis de regulação, evidenciando a profundidade com que a bioquímica estrutura e coordena processos essenciais à vida.
5.11 Lipídios e adaptação biológica #
A composição lipídica das células não é fixa; ela responde ao ambiente e, em muitos casos, determina a capacidade do organismo de sobreviver sob condições adversas. Entre os fatores mais críticos, a temperatura exerce pressão seletiva direta sobre a organização das membranas. Para manter a funcionalidade — permeabilidade seletiva, atividade enzimática e integridade estrutural — os sistemas biológicos ajustam a fluidez da membrana por meio de mudanças na composição dos lipídios, um processo conhecido como adaptação homeoviscosa [Figura].
Em ambientes frios, a tendência natural das bicamadas é aumentar a ordem das cadeias hidrocarbonadas, reduzindo a mobilidade e comprometendo funções dependentes de fluidez. Para compensar esse efeito, muitos organismos aumentam a proporção de ácidos graxos insaturados e, em alguns casos, reduzem o comprimento das cadeias. As insaturações na configuração cis introduzem “dobras” que dificultam o empacotamento, mantendo a fase líquida cristalina em temperaturas mais baixas. Em contrapartida, em ambientes quentes, a elevação da temperatura pode tornar a membrana excessivamente fluida; a resposta adaptativa inclui maior proporção de cadeias saturadas e mais longas, além de ajustes no teor de esteróis, estabilizando a estrutura [Tabela].
Plantas oferecem um modelo claro dessa plasticidade. Tecidos expostos a variações térmicas sazonais modulam o perfil de ácidos graxos dos fosfolipídios de membrana, preservando processos como fotossíntese e transporte de solutos. Em condições de estresse térmico, a manutenção da fluidez adequada é determinante para a eficiência dos complexos de membrana, incluindo transportadores e proteínas do sistema fotossintético [Figura]. Em ambientes do semiárido, além da temperatura, a disponibilidade hídrica impõe um segundo eixo de adaptação: a redução de perdas de água e a preservação da integridade de membranas sob desidratação.
Nesse contexto, a composição lipídica de membranas plasmáticas e de organelas torna-se um elemento estratégico. Aumentos relativos de lipídios que favorecem maior coesão da bicamada podem reduzir a permeabilidade e ajudar na retenção de água. Paralelamente, a presença de esteróis e esfingolipídios contribui para a formação de microdomínios mais ordenados, que estabilizam proteínas-chave envolvidas em resposta ao estresse. Em plantas tolerantes à seca, observa-se frequentemente uma reorganização da membrana que preserva a atividade de bombas e canais, mantendo gradientes iônicos essenciais mesmo sob condições de baixa disponibilidade hídrica.
Além da temperatura e da água, outros fatores ambientais influenciam a adaptação lipídica. A salinidade, por exemplo, altera o balanço osmótico e pode impactar diretamente a estrutura da membrana. Organismos expostos a ambientes salinos frequentemente ajustam a composição lipídica para preservar a integridade da bicamada e a funcionalidade de proteínas associadas. De forma semelhante, mudanças na disponibilidade de oxigênio e nutrientes podem modular a síntese e a remodelação de lipídios, integrando o estado metabólico à organização estrutural.
A adaptação lipídica não se limita à membrana plasmática. Organelas como mitocôndrias e cloroplastos apresentam composições específicas que refletem suas funções. Nos cloroplastos, por exemplo, a organização dos tilacoides depende de lipídios especializados que garantem a fluidez adequada para a dinâmica dos complexos fotossintéticos. Alterações nessas composições, induzidas por estresse ambiental, podem comprometer a eficiência da conversão de energia luminosa, impactando diretamente a produtividade vegetal.
Do ponto de vista molecular, essas respostas envolvem regulação fina de enzimas responsáveis pela síntese e modificação de lipídios, incluindo dessaturases e elongases. A ativação ou repressão dessas vias permite ajustes rápidos e direcionados na composição lipídica, conectando sinais ambientais à resposta estrutural. Esse nível de controle evidencia que a adaptação não é passiva, mas resultado de um sistema regulatório integrado que atua sobre a química das membranas.
Em termos aplicados, compreender a adaptação lipídica tem implicações diretas para a agricultura, especialmente em regiões sujeitas a estresses ambientais intensos. A seleção ou engenharia de plantas com perfis lipídicos mais resilientes pode resultar em maior tolerância a temperaturas extremas e déficit hídrico, contribuindo para a estabilidade produtiva em condições adversas.
Assim, os lipídios deixam de ser vistos apenas como componentes estruturais e passam a ser reconhecidos como elementos ativos na interface entre o organismo e o ambiente. A capacidade de modular sua composição e organização permite que sistemas biológicos mantenham a funcionalidade sob condições variáveis, evidenciando a centralidade dos lipídios na adaptação e na sobrevivência.
5.12 Interações lipídio–proteína em sistemas biológicos #
A funcionalidade das membranas biológicas não pode ser compreendida apenas a partir da organização dos lipídios ou da presença de proteínas isoladamente. O que define, de fato, o comportamento desses sistemas é a interação cooperativa entre lipídios e proteínas, formando uma unidade funcional integrada. Nesse contexto, as proteínas de membrana não operam em um ambiente neutro, mas em um meio lipídico que modula sua estrutura, dinâmica e atividade [Figura].
As proteínas de membrana podem ser classificadas, de forma geral, em integrais e periféricas. As proteínas integrais estão inseridas na bicamada lipídica, frequentemente atravessando-a por meio de domínios hidrofóbicos que interagem diretamente com as cadeias de ácidos graxos dos lipídios. Já as proteínas periféricas associam-se à superfície da membrana por interações eletrostáticas ou por ligação a lipídios específicos. Em ambos os casos, a interação com o ambiente lipídico é essencial para a estabilidade e função dessas proteínas.
Do ponto de vista estrutural, a inserção de proteínas integrais na membrana depende da compatibilidade entre seus domínios hidrofóbicos e a espessura da bicamada lipídica. Alterações na composição dos lipídios — como mudanças no comprimento das cadeias ou no grau de insaturação — podem modificar essa espessura, influenciando diretamente a conformação das proteínas. Esse fenômeno, conhecido como “acoplamento hidrofóbico”, evidencia que a estrutura proteica é sensível ao ambiente lipídico em que está inserida.
Além disso, lipídios específicos podem atuar como cofatores estruturais ou regulatórios para proteínas de membrana. Certos lipídios ligam-se de forma direta e específica a regiões da proteína, estabilizando conformações ativas ou inativas. Em outros casos, a presença de determinados lipídios altera as propriedades físicas da membrana — como fluidez, curvatura ou carga —, influenciando indiretamente a atividade proteica. Assim, a função de muitas proteínas não depende apenas de sua sequência de aminoácidos, mas também do contexto lipídico em que se encontram.
A organização lateral da membrana também desempenha papel importante nessas interações. Como discutido anteriormente, a bicamada não é homogênea, podendo apresentar microdomínios enriquecidos em certos tipos de lipídios, como esfingolipídios e colesterol. Esses domínios, frequentemente chamados de “rafts lipídicos”, funcionam como plataformas organizadoras onde proteínas específicas são concentradas. Essa organização espacial facilita interações proteína–proteína e a formação de complexos multiproteicos envolvidos em processos como sinalização celular e transporte [Figura].
Outro aspecto relevante é a influência da curvatura da membrana nas interações lipídio–proteína. Certas proteínas possuem domínios especializados capazes de reconhecer ou induzir curvaturas específicas na bicamada. Essa capacidade é essencial em processos como endocitose, exocitose e formação de vesículas, nos quais a membrana precisa ser deformada de maneira controlada. Nesses casos, a composição lipídica — incluindo a presença de lipídios com geometrias moleculares distintas — contribui para a geração e estabilização dessas curvaturas.
As interações lipídio–proteína também são fundamentais para o funcionamento de sistemas de transporte. Canais iônicos, transportadores e bombas dependem de um ambiente lipídico adequado para manter sua conformação funcional. Alterações na fluidez ou na composição da membrana podem afetar a cinética desses sistemas, influenciando diretamente o fluxo de íons e moléculas através da membrana.
Do ponto de vista dinâmico, essas interações são altamente reguláveis. Mudanças na composição lipídica, induzidas por sinais celulares ou condições ambientais, podem modificar rapidamente o comportamento das proteínas associadas. Isso permite que a célula ajuste suas funções de maneira eficiente, integrando informações externas e internas em respostas coordenadas.
Em termos fisiológicos e patológicos, a importância dessas interações é evidente. Disfunções na composição lipídica da membrana podem comprometer a atividade de proteínas essenciais, contribuindo para o desenvolvimento de diversas doenças, incluindo distúrbios neurológicos, metabólicos e cardiovasculares. Por outro lado, a modulação dessas interações representa uma estratégia promissora para intervenções terapêuticas.
Assim, as interações entre lipídios e proteínas revelam um nível de organização bioquímica no qual a função emerge da cooperação entre diferentes classes de biomoléculas. A membrana deixa de ser vista como uma simples matriz estrutural e passa a ser compreendida como um sistema dinâmico e integrado, no qual lipídios e proteínas atuam conjuntamente para sustentar a complexidade da vida celular.
5.13 Lipídios e doenças metabólicas #
A mesma eficiência bioquímica que torna os lipídios essenciais para a vida — armazenamento energético compacto, organização de membranas e participação em sinalização — pode, sob condições de desregulação, contribuir para o desenvolvimento de doenças metabólicas. O ponto central não é a presença de lipídios em si, mas o desequilíbrio entre sua síntese, transporte, armazenamento e utilização. Quando esses fluxos deixam de estar coordenados, surgem estados patológicos que refletem, em última análise, falhas na homeostase lipídica [Figura].
Um dos exemplos mais relevantes é a aterosclerose, uma condição caracterizada pelo acúmulo progressivo de lipídios, especialmente colesterol, nas paredes das artérias. Esse processo está intimamente associado às lipoproteínas de baixa densidade (LDL), que transportam colesterol para os tecidos. Em concentrações elevadas ou em formas modificadas, as LDL podem ser retidas na parede vascular, onde sofrem oxidação e desencadeiam respostas inflamatórias. Macrófagos recrutados ao local internalizam essas partículas lipídicas, transformando-se em células espumosas e contribuindo para a formação de placas ateroscleróticas [Figura].
Esse processo não é apenas um fenômeno de deposição passiva, mas uma resposta inflamatória crônica que envolve múltiplos mediadores celulares e moleculares. Com o tempo, as placas podem comprometer o fluxo sanguíneo ou tornar-se instáveis, levando a eventos agudos como infarto do miocárdio e acidente vascular cerebral. Assim, a aterosclerose exemplifica como a disfunção no transporte e no metabolismo do colesterol pode ter consequências sistêmicas graves.
Outro eixo importante das doenças lipídicas está relacionado ao acúmulo excessivo de triacilgliceróis no tecido adiposo, caracterizando a obesidade. Embora o armazenamento de lipídios seja fisiologicamente necessário, o excesso crônico de energia leva à expansão do tecido adiposo além de sua capacidade funcional. Esse estado é acompanhado por alterações na secreção de moléculas sinalizadoras, como adipocinas, e pela instalação de um ambiente inflamatório de baixo grau [Figura].
A obesidade não deve ser entendida apenas como um aumento quantitativo de gordura, mas como uma condição metabólica complexa que afeta a função de múltiplos tecidos. Um dos desdobramentos mais relevantes é a resistência à insulina, na qual células de tecidos como músculo e fígado tornam-se menos responsivas a esse hormônio. Lipídios acumulados em locais não especializados, como fígado e músculo — fenômeno conhecido como lipotoxicidade — interferem em vias de sinalização, prejudicando a captação e utilização de glicose.
Essa interconexão entre metabolismo lipídico e glicídico está na base de doenças como o diabetes mellitus tipo 2. Nesse contexto, o excesso de ácidos graxos circulantes e o acúmulo de intermediários lipídicos alteram a sinalização intracelular, contribuindo para a disfunção metabólica sistêmica. Assim, os lipídios deixam de ser apenas fontes de energia e passam a atuar como moduladores negativos do metabolismo quando presentes em excesso ou em locais inadequados.
As dislipidemias representam outro grupo importante de alterações, caracterizadas por níveis anormais de lipídios no sangue, incluindo colesterol e triacilgliceróis. Essas condições podem ter origem genética ou resultar de fatores ambientais, como dieta e estilo de vida. Independentemente da causa, as dislipidemias refletem desequilíbrios nos sistemas de transporte e regulação lipídica, frequentemente associados a maior risco cardiovascular.
Do ponto de vista molecular, muitas dessas condições envolvem alterações em enzimas-chave, receptores ou proteínas de transporte. Defeitos em receptores de LDL, por exemplo, comprometem a remoção de colesterol da circulação, enquanto alterações em enzimas que regulam o metabolismo de ácidos graxos podem levar ao acúmulo de intermediários tóxicos. Esses exemplos reforçam que a homeostase lipídica depende de uma rede complexa de interações bioquímicas.
Além das doenças cardiovasculares e metabólicas clássicas, alterações no metabolismo lipídico também estão associadas a condições neurodegenerativas, inflamatórias e até mesmo ao câncer. Lipídios participam da organização de membranas neuronais, da sinalização celular e da regulação do crescimento celular, de modo que sua desregulação pode impactar diversos sistemas biológicos.
Em síntese, os lipídios ocupam uma posição ambígua na biologia: são essenciais para a vida, mas, quando desregulados, tornam-se centrais na fisiopatologia de várias doenças. A compreensão desses processos exige uma visão integrada, na qual estrutura, função e metabolismo são considerados de forma conjunta. Esse entendimento não apenas esclarece os mecanismos das doenças, mas também abre caminhos para estratégias de prevenção e intervenção terapêutica, reforçando o papel da bioquímica como base para a medicina e a saúde.
5.14 Aplicações biotecnológicas e industriais dos lipídios #
A relevância dos lipídios ultrapassa o domínio da biologia celular e da fisiologia, estendendo-se de forma estratégica para aplicações biotecnológicas e industriais. Essa transição do laboratório para o setor produtivo é possível porque as propriedades fundamentais dos lipídios — hidrofobicidade, alta densidade energética, capacidade de auto-organização e diversidade estrutural — podem ser exploradas de maneira direcionada em diferentes contextos tecnológicos. Nesse cenário, os lipídios deixam de ser apenas objetos de estudo e passam a ser recursos funcionais para geração de valor [Figura].
Uma das aplicações mais consolidadas encontra-se na produção de biocombustíveis, especialmente o biodiesel. Esse combustível é obtido a partir de triacilgliceróis presentes em óleos vegetais ou gorduras animais, por meio de reações de transesterificação que geram ésteres de ácidos graxos. A eficiência desse processo está diretamente relacionada à estrutura dos lipídios utilizados, incluindo o grau de insaturação e o comprimento das cadeias carbônicas. Além de representar uma alternativa renovável aos combustíveis fósseis, o biodiesel conecta diretamente a bioquímica dos lipídios à sustentabilidade energética e à economia agrícola [Figura].
Na indústria alimentícia, os lipídios desempenham funções que vão muito além do fornecimento calórico. Eles são determinantes na textura, estabilidade, sabor e aparência de alimentos. Processos como hidrogenação e interesterificação são utilizados para modificar propriedades físicas dos lipídios, ajustando características como ponto de fusão e consistência. No entanto, essas modificações também levantam questões de saúde, como a formação de ácidos graxos trans, evidenciando a necessidade de equilíbrio entre funcionalidade industrial e impacto nutricional.
Outro campo de aplicação relevante é a nanotecnologia, particularmente no desenvolvimento de sistemas de liberação controlada. Lipídios anfipáticos são utilizados para formar lipossomas e nanopartículas lipídicas, estruturas capazes de encapsular moléculas bioativas, protegendo-as da degradação e direcionando sua entrega a tecidos específicos. Esse princípio tem sido amplamente explorado na farmacologia, incluindo o desenvolvimento de sistemas avançados de entrega de fármacos e vacinas, nos quais a biocompatibilidade dos lipídios representa uma vantagem significativa [Figura].
Na área cosmética, os lipídios são utilizados tanto como agentes estruturais quanto funcionais. Cremes, emulsões e sistemas de hidratação dependem da capacidade dos lipídios de formar barreiras protetoras e de interagir com a pele. A semelhança estrutural entre certos lipídios industriais e os lipídios naturais da epiderme permite o desenvolvimento de produtos que restauram a integridade da barreira cutânea, evidenciando a aplicação direta de conceitos bioquímicos em produtos de uso cotidiano.
A biotecnologia moderna também explora lipídios como alvos e ferramentas em processos industriais. Microrganismos geneticamente modificados podem ser utilizados para produzir lipídios específicos, como ácidos graxos de cadeia longa ou compostos bioativos, ampliando o espectro de aplicações. Em sistemas agrícolas, o conhecimento da bioquímica lipídica tem sido utilizado para desenvolver plantas com perfis lipídicos otimizados, seja para fins nutricionais, seja para aplicações industriais.
Além disso, lipídios desempenham papel importante em processos ambientais, como biorremediação. Certos microrganismos utilizam compostos lipídicos como fonte de carbono, sendo capazes de degradar poluentes orgânicos hidrofóbicos. Essa capacidade pode ser explorada para mitigar impactos ambientais, especialmente em áreas contaminadas por hidrocarbonetos.
No contexto do semiárido e da produção agrícola, a utilização estratégica de lipídios também pode representar uma oportunidade de inovação. Culturas adaptadas a condições de estresse hídrico frequentemente apresentam perfis lipídicos específicos que podem ser explorados tanto para resistência ambiental quanto para aplicações industriais. Isso abre espaço para integração entre bioquímica, agronomia e desenvolvimento regional.
Do ponto de vista econômico, os lipídios representam uma interface entre ciência básica e mercado, com cadeias produtivas que vão desde a produção primária até aplicações de alto valor agregado. A capacidade de manipular suas propriedades químicas e físicas permite a criação de produtos com características específicas, adaptadas a diferentes demandas.
Assim, as aplicações biotecnológicas e industriais dos lipídios demonstram como princípios fundamentais da bioquímica podem ser traduzidos em soluções práticas e economicamente relevantes. Ao conectar estrutura molecular, função biológica e inovação tecnológica, os lipídios consolidam-se como uma das classes de biomoléculas mais versáteis, com impacto direto em setores estratégicos como energia, saúde, alimentação e meio ambiente.
5.15 Integração conceitual: lipídios como eixo estrutural e funcional #
Ao longo deste capítulo, os lipídios foram apresentados sob diferentes perspectivas — estrutural, físico-química, funcional e aplicada. No entanto, a compreensão plena de sua relevância em sistemas biológicos exige uma visão integrada, na qual essas dimensões não são tratadas de forma isolada, mas como partes de um mesmo sistema dinâmico. Os lipídios ocupam uma posição estratégica na bioquímica exatamente por conectarem, de maneira contínua, organização molecular, estrutura celular e metabolismo energético.
A primeira dimensão dessa integração está na relação entre estrutura química e organização supramolecular. A natureza anfipática de muitos lipídios não apenas determina sua interação com a água, mas conduz diretamente à formação de membranas biológicas. Essas estruturas, por sua vez, criam compartimentos, estabelecem gradientes e permitem a especialização funcional das células. Assim, uma propriedade molecular simples — a coexistência de regiões hidrofóbicas e hidrofílicas — torna-se o fundamento da organização da vida celular [Figura].
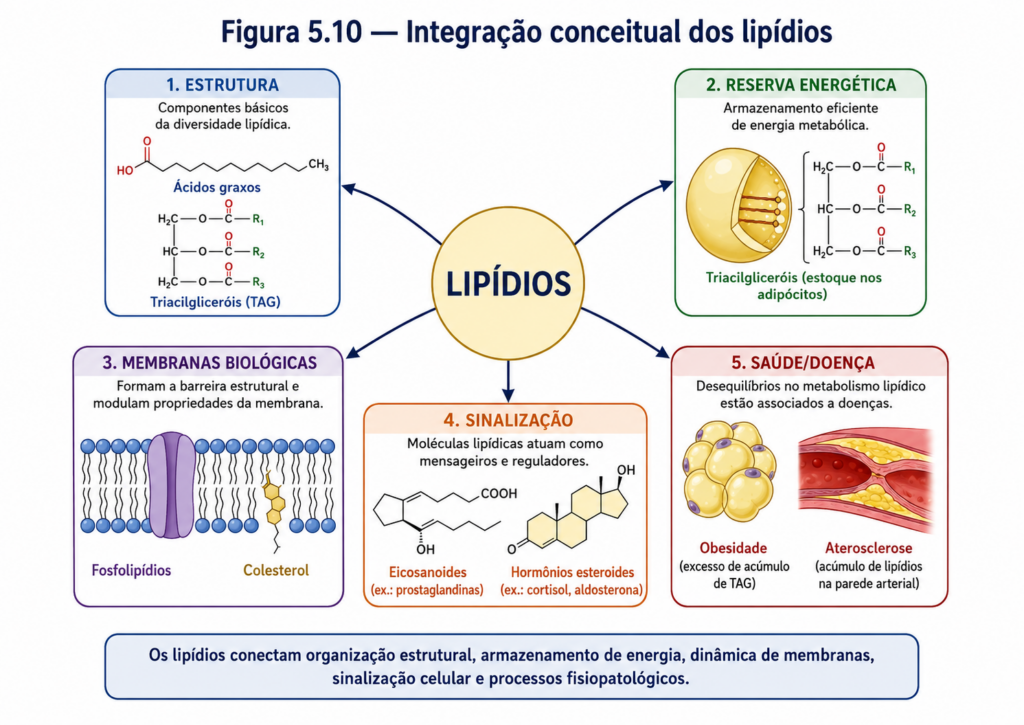
Em paralelo, os lipídios conectam estrutura e energia de maneira singular. Enquanto carboidratos fornecem energia de acesso rápido, os lipídios atuam como reservas de longo prazo, armazenando grandes quantidades de energia em forma compacta. Essa função energética não está dissociada de sua organização estrutural: os mesmos ácidos graxos que compõem triacilgliceróis participam da formação de membranas e de moléculas sinalizadoras. Dessa forma, os lipídios transitam continuamente entre papéis estruturais e metabólicos, dependendo das demandas do organismo.
A integração também se manifesta na interface com outras classes de biomoléculas. Proteínas de membrana dependem do ambiente lipídico para manter sua conformação e atividade, enquanto carboidratos associados a lipídios participam de processos de reconhecimento celular. Além disso, lipídios sinalizadores conectam o estado metabólico da célula a respostas regulatórias, influenciando vias de sinalização e expressão gênica. Essa interdependência evidencia que os lipídios não atuam isoladamente, mas como componentes de redes bioquímicas complexas.
Outro eixo integrador importante é a relação entre lipídios e adaptação biológica. A capacidade de modificar a composição lipídica em resposta a mudanças ambientais — como temperatura, disponibilidade hídrica e condições nutricionais — permite que organismos mantenham a funcionalidade celular sob diferentes contextos. Esse ajuste fino conecta diretamente a química das moléculas ao desempenho fisiológico e à sobrevivência do organismo.
Do ponto de vista sistêmico, os lipídios participam de fluxos metabólicos que envolvem múltiplos órgãos e tecidos. A digestão, absorção e transporte por lipoproteínas garantem sua distribuição eficiente, enquanto mecanismos regulatórios controlam seu armazenamento e mobilização. Alterações nesses fluxos podem resultar em estados patológicos, como obesidade, dislipidemias e doenças cardiovasculares, evidenciando que a integração lipídica é essencial para a homeostase.
Essa visão integrada prepara o terreno para a próxima etapa do estudo da bioquímica: o metabolismo. Até aqui, os lipídios foram analisados principalmente em termos de estrutura e função. No entanto, sua verdadeira dinâmica emerge quando se considera como são sintetizados, degradados e interconvertidos em diferentes contextos fisiológicos. A partir desse ponto, o foco desloca-se para as vias metabólicas que governam o fluxo de energia e matéria, nas quais os lipídios desempenham papel central.
Em síntese, os lipídios devem ser compreendidos como um eixo estruturante da bioquímica, capaz de conectar níveis distintos de organização biológica. Desde a química das ligações carbono–hidrogênio até a complexidade dos sistemas celulares e organismais, eles atuam como mediadores entre estrutura, energia e informação. Essa integração não apenas explica sua ubiquidade nos sistemas vivos, mas também reforça sua importância como base para o entendimento da fisiologia e das aplicações biotecnológicas.
Com isso, encerra-se a abordagem estrutural dos lipídios e abre-se o caminho para a análise de sua dinâmica metabólica, onde os princípios aqui estabelecidos serão mobilizados para compreender como a vida transforma, regula e utiliza essas moléculas em escala sistêmica.
Referencias #
NELSON, David L.; COX, Michael M. Princípios de bioquímica de Lehninger. 6. ed. Porto Alegre: Artmed, 2014.
VOET, Donald; VOET, Judith G.; PRATT, Charlotte W. Fundamentos de bioquímica: a vida em nível molecular. 4. ed. Porto Alegre: Artmed, 2014.
STRYER, Lubert. Bioquímica. 7. ed. Rio de Janeiro: Guanabara Koogan, 2014.
MURRAY, Robert K. et al. Harper: bioquímica ilustrada. 31. ed. Porto Alegre: AMGH, 2018.
Perguntas de estudo dirigido — Capítulo 5: Lipídios #
- Explique por que os lipídios não são definidos por uma única estrutura química, mas por propriedades físico-químicas comuns, especialmente a baixa solubilidade em água.
- Compare ácidos graxos saturados e insaturados cis, relacionando suas diferenças estruturais com o ponto de fusão, o empacotamento molecular e a fluidez das membranas.
- Descreva a formação dos triacilgliceróis e explique por que eles são considerados moléculas altamente eficientes para armazenamento energético.
- Explique como fosfolipídios e esfingolipídios se organizam em meio aquoso e por que essa propriedade é essencial para a formação das membranas biológicas.
- Analise o papel do colesterol na membrana celular, destacando seu efeito sobre a fluidez em temperaturas altas e baixas, e sua importância como precursor de moléculas biologicamente ativas.
Pergunta para estimular pesquisa #
Como alterações na composição lipídica das membranas celulares podem influenciar a tolerância de plantas a estresses ambientais, como seca, salinidade e altas temperaturas, especialmente em sistemas agrícolas do semiárido?



