Os carboidratos são fundamentais como fonte de energia e componentes estruturais. Este capítulo aborda sua diversidade estrutural, desde monossacarídeos até polissacarídeos complexos, destacando isomeria e ligações glicosídicas. A relação entre estrutura e função é explorada em contextos biológicos, evidenciando diferenças entre sistemas vegetais e animais. A compreensão desses compostos é essencial para interpretar processos metabólicos energéticos.
4.1 Introdução aos carboidratos na bioquímica #
Ao observar qualquer sistema biológico — de uma folha submetida à intensa radiação solar no semiárido até o tecido muscular humano em atividade — torna-se evidente que a vida depende de um fluxo contínuo de energia e de estruturas capazes de armazená-la, transformá-la e utilizá-la com precisão. Nesse contexto, os carboidratos emergem como uma das classes mais fundamentais de biomoléculas, atuando simultaneamente como fonte primária de energia, reserva metabólica e elemento estrutural de organismos vivos.
Quimicamente, os carboidratos são compostos orgânicos constituídos predominantemente por carbono, hidrogênio e oxigênio, frequentemente descritos pela fórmula empírica geral (CH₂O)ₙ. Essa representação, embora simplificada, mascara uma diversidade estrutural notável, que se manifesta desde pequenas moléculas altamente reativas até polímeros complexos com funções especializadas. A chave para compreender essa diversidade reside na arquitetura molecular desses compostos, cuja organização determina não apenas suas propriedades químicas, mas também seu papel biológico.
Do ponto de vista evolutivo, os carboidratos ocupam uma posição central na bioquímica da vida. A fotossíntese, processo que sustenta a maior parte da biosfera, converte energia luminosa em energia química armazenada na forma de açúcares, principalmente glicose. Esse processo estabelece uma ponte direta entre o fluxo de energia solar e a organização molecular da matéria viva, permitindo que organismos heterotróficos utilizem esses compostos como combustível metabólico. Assim, os carboidratos não são apenas moléculas funcionais; são, na prática, o principal elo entre energia ambiental e metabolismo celular.
No metabolismo celular, a glicose desempenha papel central como substrato energético. Sua oxidação controlada, por meio de vias como a glicólise e o ciclo do ácido cítrico, resulta na produção de ATP, a moeda energética universal das células. Esse processo ilustra um princípio fundamental da bioquímica: a extração gradual e eficiente de energia a partir de ligações químicas, permitindo que sistemas biológicos realizem trabalho sem violar as leis da termodinâmica. [Equação]
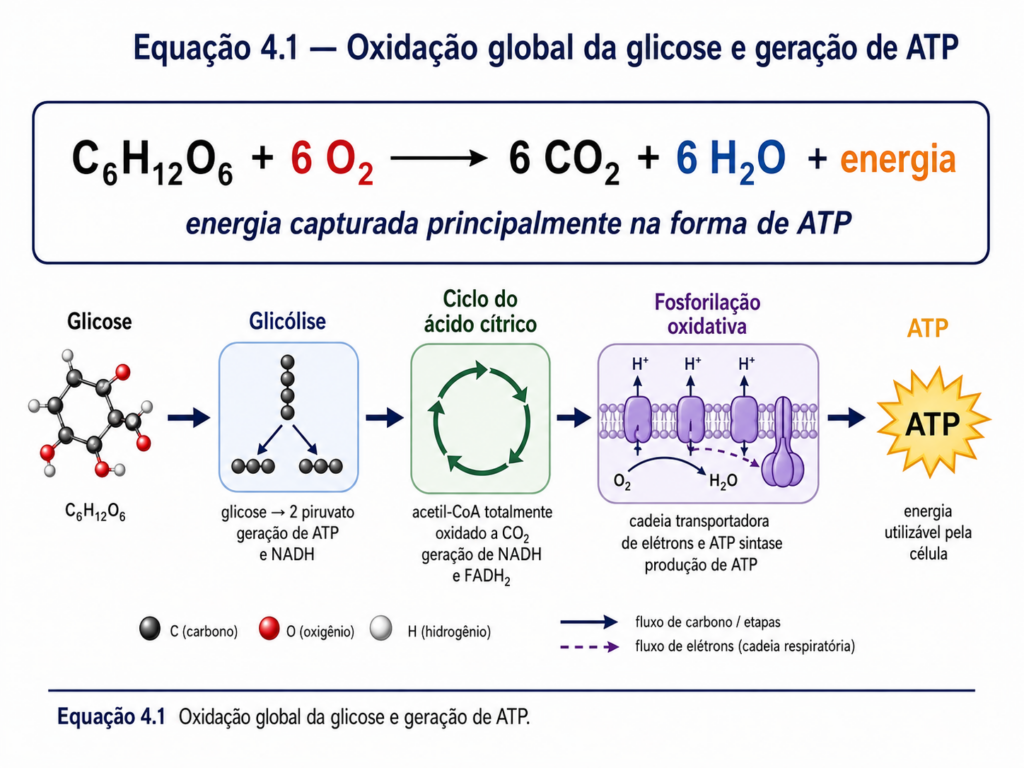
Além da função energética, os carboidratos exercem papel estrutural indispensável. Polímeros como a celulose, principal componente da parede celular vegetal, conferem rigidez e resistência mecânica, permitindo que plantas mantenham sua arquitetura mesmo sob condições ambientais adversas. Em organismos animais, estruturas glicídicas associadas a proteínas e lipídios participam da formação de matrizes extracelulares e superfícies celulares, influenciando diretamente processos como adesão, comunicação e reconhecimento molecular. [Figura]
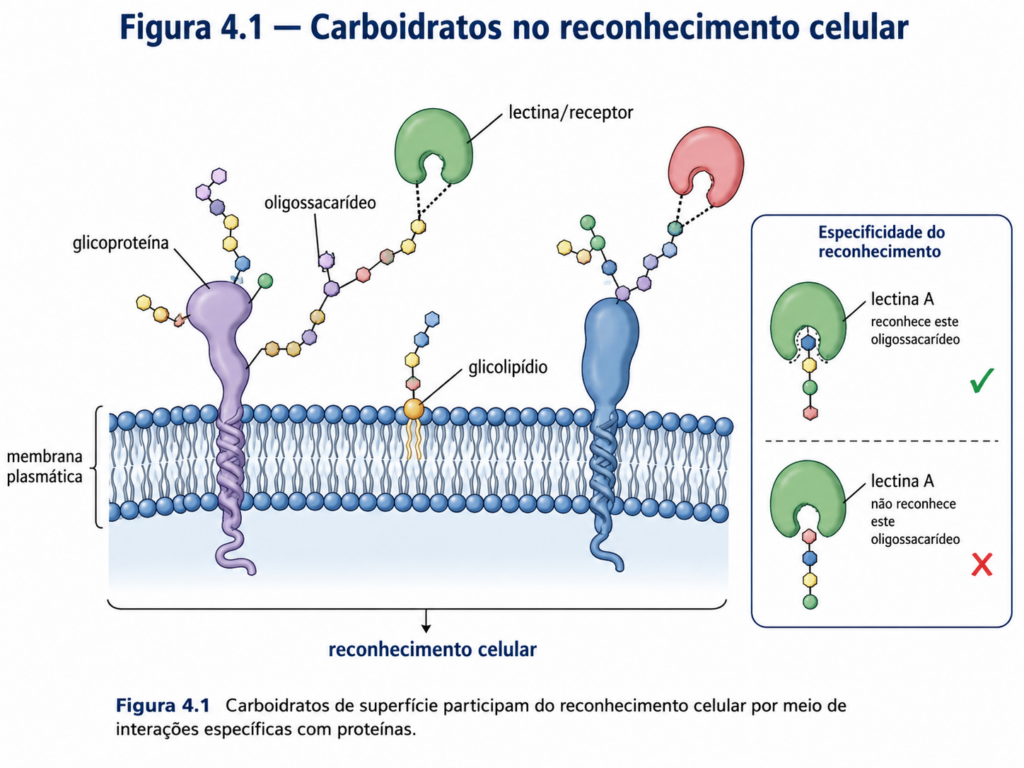
Outro aspecto crítico dos carboidratos é sua participação em sistemas de informação biológica. Embora não armazenem informação genética como os ácidos nucleicos, oligossacarídeos presentes na superfície celular atuam como marcadores moleculares altamente específicos. Esses “códigos glicídicos” são reconhecidos por proteínas específicas, modulando interações celulares, respostas imunológicas e processos de sinalização. Essa função evidencia que os carboidratos transcendem o papel de meros combustíveis metabólicos, integrando-se a redes complexas de comunicação molecular.
A relevância dos carboidratos torna-se ainda mais evidente quando se analisam suas implicações fisiológicas e patológicas. Alterações no metabolismo da glicose estão diretamente associadas a condições como diabetes mellitus, caracterizada por falhas na regulação da captação e utilização desse açúcar. Da mesma forma, a estrutura e digestibilidade de polissacarídeos influenciam processos nutricionais, microbiota intestinal e desempenho produtivo em sistemas agrícolas, reforçando a importância desses compostos tanto na saúde humana quanto na produção de alimentos.
Do ponto de vista químico, os carboidratos também exemplificam de forma clara a relação entre estrutura e função, um dos pilares da bioquímica moderna. Pequenas variações na configuração espacial de grupos funcionais podem resultar em diferenças significativas na reatividade e no reconhecimento enzimático dessas moléculas. Essa sensibilidade estrutural é explorada por sistemas biológicos para garantir especificidade e eficiência em processos metabólicos e regulatórios.
Portanto, o estudo dos carboidratos não deve ser encarado como um tópico isolado dentro da bioquímica, mas como um eixo integrador que conecta energia, estrutura e informação. Ao longo deste capítulo, serão explorados os fundamentos químicos que governam essas moléculas, suas variações estruturais e suas funções biológicas, estabelecendo a base necessária para a compreensão das vias metabólicas que sustentam a vida.
Essa abordagem permitirá avançar além da memorização de estruturas e nomenclaturas, conduzindo a uma compreensão mecanística e aplicada dos carboidratos — exatamente o nível de domínio exigido para interpretar fenômenos biológicos, desenvolver soluções tecnológicas e tomar decisões estratégicas em contextos científicos, agronômicos e industriais.
4.2 Classificação dos carboidratos #
A diversidade funcional dos carboidratos decorre diretamente de sua organização estrutural. Embora compartilhem uma base química comum — a presença de grupos carbonila e hidroxila em cadeias de carbono —, essas moléculas se distribuem em diferentes níveis de complexidade, cuja classificação não é meramente didática, mas reflete propriedades químicas, comportamentos metabólicos e funções biológicas distintas.
A forma mais fundamental de organização dos carboidratos é representada pelos monossacarídeos, unidades básicas que não podem ser hidrolisadas em moléculas menores. Essas estruturas simples constituem o ponto de partida para toda a arquitetura glicídica nos sistemas biológicos. A glicose, por exemplo, destaca-se como o principal combustível metabólico das células, sendo rapidamente mobilizada e convertida em energia química. Outros monossacarídeos, como frutose e galactose, participam de rotas metabólicas específicas, evidenciando que mesmo pequenas variações estruturais resultam em destinos bioquímicos distintos.
À medida que monossacarídeos se unem por meio de ligações glicosídicas, surgem os oligossacarídeos, compostos por um número limitado de unidades (tipicamente entre duas e dez). Os dissacarídeos — como sacarose, lactose e maltose — são exemplos clássicos dessa categoria e desempenham papel relevante na nutrição, atuando como formas de transporte e armazenamento temporário de energia. No entanto, a importância dos oligossacarídeos vai além do metabolismo energético. Quando ligados a proteínas e lipídios, formam estruturas complexas envolvidas em reconhecimento celular, adesão e sinalização, funcionando como elementos-chave em sistemas de comunicação molecular. [Figura]
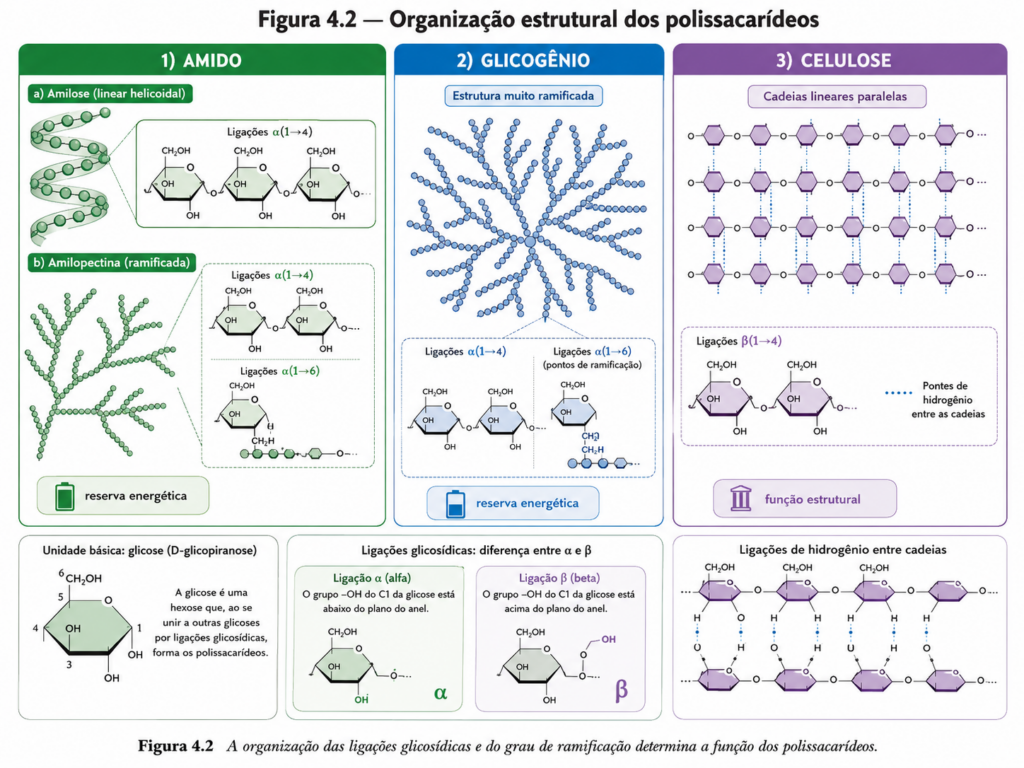
O nível mais elevado de organização estrutural é alcançado pelos polissacarídeos, macromoléculas formadas por longas cadeias de monossacarídeos. Aqui, a função biológica passa a ser fortemente determinada pela natureza das ligações glicosídicas e pelo grau de ramificação das cadeias. Polissacarídeos de reserva, como o glicogênio em animais e o amido em plantas, são altamente organizados para permitir mobilização rápida de energia. Já os polissacarídeos estruturais, como a celulose, apresentam arranjos lineares e altamente estáveis, conferindo resistência mecânica e suporte às células vegetais. Essa distinção ilustra um princípio central: a mesma unidade básica — a glicose — pode originar materiais com propriedades completamente distintas dependendo de sua organização tridimensional. [Figura]
Além dessa classificação baseada no número de unidades monoméricas, os carboidratos também podem ser categorizados segundo sua composição química. Nesse contexto, distinguem-se os homopolissacarídeos, constituídos por um único tipo de monossacarídeo, e os heteropolissacarídeos, formados por diferentes unidades. Essa diferenciação é particularmente relevante em sistemas biológicos complexos, como a matriz extracelular, onde a combinação de diferentes açúcares gera estruturas com propriedades físico-químicas específicas, como viscosidade, retenção de água e capacidade de interação com proteínas.
Outro critério importante envolve a natureza funcional dos grupos químicos presentes. Carboidratos podem ser classificados como aldoses ou cetoses, dependendo da posição do grupo carbonila. Essa distinção, aparentemente sutil, tem implicações diretas na reatividade química e no metabolismo dessas moléculas. Paralelamente, a classificação pelo número de carbonos — trioses, tetroses, pentoses e hexoses — fornece uma base sistemática para entender a diversidade estrutural e suas implicações biológicas.
Do ponto de vista bioquímico, essa organização hierárquica — do monossacarídeo ao polissacarídeo — não é apenas uma questão de complexidade crescente, mas um reflexo direto da estratégia evolutiva dos sistemas vivos. Estruturas simples são utilizadas para reações rápidas e fluxos metabólicos dinâmicos, enquanto estruturas complexas são empregadas para armazenamento, proteção e interação molecular. Esse arranjo evidencia um padrão recorrente na biologia: a modularidade, na qual unidades simples são combinadas de diferentes maneiras para gerar funções altamente especializadas.
Compreender a classificação dos carboidratos, portanto, é estabelecer um mapa conceitual que permite antecipar comportamento químico, função biológica e relevância metabólica. Essa base será essencial para os tópicos subsequentes, nos quais a análise estrutural detalhada e as propriedades químicas dessas moléculas serão exploradas com maior profundidade, conectando forma molecular e função biológica em nível mecanístico.
4.3 Estrutura química dos monossacarídeos #
A compreensão dos carboidratos em nível molecular inicia-se necessariamente pela análise dos monossacarídeos, as unidades estruturais mais simples dessa classe de biomoléculas. Embora sejam frequentemente apresentados como entidades químicas elementares, sua organização interna revela uma sofisticação estrutural que sustenta grande parte da diversidade funcional observada nos sistemas biológicos.
Quimicamente, os monossacarídeos são polihidroxialdeídos ou polihidroxicetonas, isto é, moléculas que contêm múltiplos grupos hidroxila (–OH) associados a um grupo carbonila (C=O). Essa dupla característica confere a essas moléculas uma elevada reatividade e capacidade de interação com o meio aquoso, tornando-as altamente solúveis e metabolicamente versáteis. A presença simultânea desses grupos funcionais estabelece as bases para uma ampla gama de reações bioquímicas, incluindo oxidação, redução, esterificação e formação de ligações glicosídicas.
A classificação estrutural inicial dos monossacarídeos deriva da posição do grupo carbonila na cadeia carbônica. Quando esse grupo está localizado na extremidade da molécula, o composto é denominado aldose, enquanto sua posição interna caracteriza uma cetose. Essa distinção, embora simples, tem implicações profundas na reatividade química e nas vias metabólicas em que essas moléculas participam. A glicose, por exemplo, é uma aldo-hexose amplamente utilizada como fonte energética, enquanto a frutose, uma ceto-hexose, segue rotas metabólicas parcialmente distintas, refletindo sua organização estrutural específica.
Outro critério fundamental é o número de átomos de carbono presentes na cadeia. Monossacarídeos podem variar de três a sete carbonos, sendo denominados trioses, tetroses, pentoses, hexoses e heptoses. Entre esses, as hexoses — particularmente a glicose — assumem papel central no metabolismo energético, enquanto as pentoses, como a ribose e a desoxirribose, são componentes essenciais dos ácidos nucleicos. Essa diversidade em tamanho molecular amplia o repertório funcional dos carboidratos, permitindo sua participação em processos que vão desde o armazenamento de energia até a codificação de informação genética.
Em solução aquosa, entretanto, os monossacarídeos raramente permanecem em sua forma linear. Devido à proximidade entre o grupo carbonila e uma das hidroxilas da própria molécula, ocorre uma reação intramolecular que leva à formação de estruturas cíclicas estáveis, conhecidas como hemiacetais (no caso das aldoses) ou hemicetais (no caso das cetoses). Esse processo resulta em anéis de cinco ou seis membros, denominados furanoses e piranoses, respectivamente. [Figura]
A ciclização introduz um novo elemento estrutural de grande relevância: o carbono anomérico. Esse átomo de carbono, originado a partir do carbono carbonílico, torna-se um centro quiral adicional, permitindo a existência de duas configurações distintas, denominadas anômeros α e β. Essas formas diferem na orientação espacial do grupo hidroxila ligado ao carbono anomérico e podem interconverter-se em solução aquosa por meio de um processo conhecido como mutarrotação. Esse fenômeno não é apenas uma curiosidade estrutural, mas possui implicações diretas na reatividade química e no reconhecimento enzimático dos carboidratos.
A representação dessas estruturas cíclicas pode ser realizada por diferentes modelos, sendo a projeção de Haworth amplamente utilizada para descrever a organização planar dos anéis. No entanto, para uma compreensão mais precisa da estabilidade conformacional, torna-se necessário considerar modelos tridimensionais, como a conformação em cadeira, especialmente no caso das piranoses. Nessa configuração, a distribuição espacial dos grupos substituintes influencia diretamente a estabilidade da molécula, favorecendo arranjos que minimizam repulsões estéricas e interações desfavoráveis.
Essa sensibilidade estrutural evidencia um dos princípios centrais da bioquímica: pequenas alterações na disposição espacial de átomos podem resultar em mudanças significativas nas propriedades químicas e biológicas de uma molécula. No caso dos monossacarídeos, essa característica é explorada extensivamente pelos sistemas biológicos para garantir especificidade em processos metabólicos e interações moleculares.
Portanto, a estrutura química dos monossacarídeos não deve ser interpretada apenas como uma descrição estática de ligações e grupos funcionais, mas como um sistema dinâmico, altamente organizado e adaptado às exigências do metabolismo celular. A partir dessa base estrutural, torna-se possível compreender como essas unidades simples dão origem a moléculas mais complexas e desempenham papéis essenciais na bioquímica da vida.
4.4 Estereoquímica dos carboidratos #
A estrutura dos monossacarídeos, embora aparentemente simples, revela um nível de complexidade adicional quando analisada sob a perspectiva tridimensional. Essa complexidade é consequência direta da presença de múltiplos centros quirais, átomos de carbono ligados a quatro substituintes diferentes, que conferem aos carboidratos uma rica variedade de formas espaciais. Essa diversidade não é apenas estrutural; ela é determinante para o reconhecimento molecular, a reatividade química e a função biológica dessas moléculas.
Nos monossacarídeos, quase todos os átomos de carbono — com exceção do carbono carbonílico na forma linear — são potencialmente quirais. Isso significa que uma única fórmula molecular pode originar múltiplos estereoisômeros, compostos que possuem a mesma conectividade de átomos, mas diferem na orientação espacial de seus grupos funcionais. A quantidade de estereoisômeros possíveis cresce exponencialmente com o número de carbonos quirais, o que explica a enorme diversidade estrutural observada entre os carboidratos.
A forma mais fundamental de estereoisomeria é representada pelos enantiômeros, pares de moléculas que são imagens especulares não sobreponíveis, análogas às mãos direita e esquerda. Nos carboidratos, essa distinção é expressa pela notação D e L, baseada na configuração do carbono quiral mais distante do grupo carbonila. Por convenção, a maioria dos carboidratos biologicamente ativos pertence à série D, incluindo a glicose, que ocupa posição central no metabolismo. Essa preferência não é aleatória, mas resultado de processos evolutivos que selecionaram sistemas enzimáticos altamente específicos para determinadas configurações espaciais. [Figura]
Além dos enantiômeros, os carboidratos apresentam outro tipo importante de estereoisomeria: os epímeros. Epímeros são moléculas que diferem na configuração de apenas um único carbono quiral. Um exemplo clássico é a diferença entre glicose e galactose, que varia apenas na orientação do grupo hidroxila em um carbono específico. Apesar dessa alteração aparentemente mínima, as implicações biológicas são significativas, exigindo vias metabólicas específicas para a conversão entre essas formas. Esse fato evidencia que, em sistemas biológicos, pequenas mudanças estruturais podem demandar mecanismos enzimáticos inteiros para serem compensadas.
Outro conjunto de estereoisômeros relevantes são os diastereoisômeros, que diferem em mais de um centro quiral, mas não são imagens especulares entre si. Essa categoria amplia ainda mais o repertório estrutural dos carboidratos, contribuindo para a diversidade de funções que essas moléculas desempenham.
A importância da estereoquímica torna-se ainda mais evidente quando se considera o processo de ciclização dos monossacarídeos. A formação de estruturas cíclicas introduz um novo centro quiral, o carbono anomérico, gerando os chamados anômeros α e β. Esses isômeros diferem na orientação do grupo hidroxila ligado ao carbono anomérico e apresentam propriedades químicas e biológicas distintas. Enzimas e proteínas frequentemente distinguem de forma rigorosa entre essas duas formas, o que reforça o papel crítico da estereoquímica no reconhecimento molecular.
Do ponto de vista bioquímico, essa especificidade tridimensional é essencial. Enzimas que catalisam reações envolvendo carboidratos possuem sítios ativos altamente complementares à configuração espacial de seus substratos. Assim, uma alteração na orientação de um único grupo funcional pode impedir completamente a interação com a enzima, bloqueando a reação. Esse nível de precisão é um dos fundamentos da eficiência e seletividade dos sistemas biológicos.
Além disso, a estereoquímica dos carboidratos influencia diretamente suas propriedades físicas, como solubilidade e capacidade de formar interações intermoleculares. Em polissacarídeos, por exemplo, a orientação das ligações glicosídicas — que depende da configuração estereoquímica — determina se a molécula será flexível e ramificada, como o glicogênio, ou rígida e estrutural, como a celulose. [Figura]
Portanto, a estereoquímica dos carboidratos não é um detalhe acessório, mas um elemento central para a compreensão de sua função biológica. Ela estabelece a base para a especificidade molecular que governa processos metabólicos, interações celulares e organização estrutural nos organismos vivos. Ao dominar esses princípios, torna-se possível interpretar não apenas a diversidade dos carboidratos, mas também a lógica subjacente à sua utilização nos sistemas biológicos.
4.5 Conformações cíclicas e estabilidade #
A ciclização dos monossacarídeos representa um ponto de inflexão na compreensão da estrutura dos carboidratos. Embora a forma linear seja essencial para explicar sua reatividade inicial, é na forma cíclica que essas moléculas efetivamente existem e operam nos sistemas biológicos. No entanto, essa forma cíclica não é estática; ela apresenta diferentes arranjos espaciais, denominados conformações, cuja estabilidade é determinada por princípios físico-químicos fundamentais.
Quando um monossacarídeo, como a glicose, forma um anel cíclico, a representação mais simples dessa estrutura é feita por meio da projeção de Haworth, que descreve o anel como plano. Essa abordagem, embora útil para visualização inicial, não reflete a realidade tridimensional da molécula. Na prática, os anéis — especialmente os de seis membros (piranoses) — adotam conformações espaciais que minimizam tensões internas e interações desfavoráveis entre os átomos.
A conformação mais estável para a maioria das hexoses cíclicas é a chamada conformação em cadeira, na qual os átomos do anel ocupam posições alternadas acima e abaixo de um plano imaginário. Essa disposição permite que os grupos substituintes — como hidroxilas e hidrogênios — se posicionem de forma a reduzir repulsões estéricas. Cada substituinte pode ocupar duas posições distintas: axial, perpendicular ao plano do anel, ou equatorial, aproximadamente paralela a esse plano. [Figura]
A estabilidade da molécula está diretamente relacionada à distribuição desses grupos entre posições axiais e equatoriais. Em geral, grupos volumosos, como as hidroxilas, tendem a ocupar posições equatoriais, onde há menor interferência espacial com outros substituintes. Esse arranjo reduz interações desfavoráveis, como repulsões entre grupos próximos, tornando a conformação energeticamente mais favorável.
Um exemplo clássico dessa estabilidade é observado na β-D-glicopiranose, a forma predominante da glicose em solução. Nessa conformação, praticamente todos os grupos hidroxila ocupam posições equatoriais, o que resulta em uma estrutura altamente estável. Em contraste, a forma α apresenta pelo menos um grupo hidroxila em posição axial, aumentando a repulsão estérica e, consequentemente, reduzindo sua estabilidade relativa. Essa diferença energética, embora pequena, é suficiente para influenciar a proporção das formas presentes em equilíbrio. [Figura]
Além da conformação em cadeira, existem outras possíveis, como a conformação em barco. No entanto, essas formas são significativamente menos estáveis devido ao aumento de interações estéricas e tensões angulares. Como resultado, elas ocorrem em proporções muito menores e têm relevância limitada em condições fisiológicas.
Outro aspecto importante é a dinâmica dessas conformações. Os anéis cíclicos não são estruturas rígidas; eles podem sofrer interconversões conformacionais, embora essas mudanças estejam associadas a barreiras energéticas específicas. Esse comportamento dinâmico permite que os carboidratos ajustem sua estrutura em resposta ao ambiente molecular, facilitando interações com enzimas, proteínas e outras biomoléculas.
A estabilidade conformacional dos carboidratos também tem implicações diretas em sua função biológica. Em polissacarídeos, por exemplo, a orientação espacial das unidades monoméricas influencia a arquitetura tridimensional da macromolécula. A celulose, formada por ligações β(1→4), apresenta cadeias lineares que se organizam em estruturas altamente rígidas e resistentes. Já o glicogênio, com ligações α(1→4) e ramificações α(1→6), possui uma estrutura mais compacta e acessível, adequada para armazenamento e mobilização rápida de energia. [Figura]
Essas diferenças estruturais emergem, em última análise, da combinação entre estereoquímica e conformação. Assim, compreender as conformações cíclicas dos monossacarídeos é essencial para interpretar não apenas a estabilidade dessas moléculas isoladas, mas também o comportamento de sistemas biológicos mais complexos.
Portanto, a análise das conformações cíclicas revela que a estrutura dos carboidratos é governada por princípios de minimização de energia e organização espacial. Essa base permite compreender como pequenas variações na disposição tridimensional dos átomos podem resultar em diferenças significativas na estabilidade, reatividade e função biológica dessas moléculas.
4.6 Derivados de monossacarídeos #
A versatilidade funcional dos carboidratos não se limita às formas básicas dos monossacarídeos. Em sistemas biológicos, essas moléculas raramente permanecem em sua forma “pura”. Pelo contrário, sofrem modificações químicas específicas que expandem significativamente seu repertório estrutural e funcional. Esses compostos modificados, conhecidos como derivados de monossacarídeos, desempenham papéis centrais em processos metabólicos, estruturais e regulatórios.
Essas modificações ocorrem, em geral, sobre grupos hidroxila ou sobre o grupo carbonila, resultando em novas propriedades químicas e novas possibilidades de interação molecular. Um dos derivados mais importantes é representado pelos açúcares fosforilados. A adição de grupos fosfato, catalisada por enzimas específicas, transforma monossacarídeos em intermediários-chave do metabolismo energético. A glicose-6-fosfato, por exemplo, constitui um ponto de convergência metabólica, participando de vias como glicólise, glicogênese e via das pentoses fosfato. A fosforilação também impede a difusão dessas moléculas através da membrana celular, mantendo-as no interior da célula e permitindo controle metabólico preciso. [Equação]
Outro grupo relevante é o dos açúcares amino, nos quais um grupo hidroxila é substituído por um grupo amino (–NH₂), frequentemente acetilado. Compostos como a glucosamina e a N-acetilglicosamina são componentes fundamentais de estruturas biológicas, incluindo glicoproteínas, glicolipídios e polissacarídeos estruturais como a quitina. Essas modificações introduzem novas propriedades químicas, como a capacidade de formar ligações adicionais e interagir com outras biomoléculas, ampliando a funcionalidade dessas estruturas.
Os ácidos urônicos constituem outro exemplo importante de derivação estrutural. Nesses compostos, o grupo hidroxila terminal é oxidado a um grupo carboxila, conferindo caráter ácido à molécula. O ácido glucurônico, por exemplo, desempenha papel essencial na detoxificação de compostos no fígado, participando de reações de conjugação que aumentam a solubilidade de substâncias hidrofóbicas, facilitando sua excreção. Esse mecanismo ilustra como modificações simples podem ser exploradas para funções fisiológicas críticas.
Além disso, existem os desoxiaçúcares, nos quais um ou mais grupos hidroxila são substituídos por hidrogênio. A desoxirribose, componente estrutural do DNA, é um exemplo clássico. Essa modificação específica confere estabilidade adicional à molécula de DNA, diferenciando-a quimicamente da ribose presente no RNA. Essa distinção estrutural está diretamente relacionada às funções biológicas dessas moléculas, particularmente no armazenamento e na transmissão da informação genética.
Outro conjunto relevante de derivados inclui os álcoois de açúcar (ou polióis), formados pela redução do grupo carbonila a um grupo hidroxila adicional. Compostos como o sorbitol e o manitol desempenham funções osmóticas e podem atuar como intermediários metabólicos. Em determinadas condições fisiológicas, o acúmulo desses compostos pode estar associado a processos patológicos, como ocorre em complicações do diabetes, evidenciando novamente a importância do controle metabólico dessas transformações.
Essas modificações químicas não ocorrem de forma aleatória; são altamente reguladas e catalisadas por enzimas específicas, refletindo a precisão dos sistemas bioquímicos. A introdução ou remoção de grupos funcionais altera propriedades como carga, polaridade e reatividade, permitindo que os derivados de monossacarídeos desempenhem funções especializadas em diferentes contextos celulares.
Do ponto de vista sistêmico, os derivados de monossacarídeos funcionam como pontos de interseção entre diferentes vias metabólicas. Eles conectam o metabolismo de carboidratos a processos como síntese de macromoléculas, sinalização celular e regulação gênica. Essa integração evidencia que os carboidratos não são apenas fontes de energia, mas componentes dinâmicos de redes bioquímicas complexas.
Portanto, o estudo dos derivados de monossacarídeos amplia a compreensão da bioquímica dos carboidratos, revelando como modificações estruturais específicas são utilizadas para expandir a funcionalidade dessas moléculas. Essa capacidade de modulação química é um dos fatores que sustentam a diversidade e a eficiência dos sistemas biológicos, permitindo que estruturas simples sejam transformadas em componentes altamente especializados e indispensáveis à vida.
4.7 Ligações glicosídicas #
A formação de estruturas mais complexas a partir de monossacarídeos depende de um tipo específico de ligação covalente: a ligação glicosídica. Esse processo representa um passo fundamental na bioquímica dos carboidratos, pois transforma unidades simples em oligossacarídeos e polissacarídeos com propriedades químicas e funções biológicas amplamente distintas. Trata-se, portanto, de um mecanismo central na construção da diversidade estrutural dos carboidratos.
Do ponto de vista químico, a ligação glicosídica resulta de uma reação de condensação entre o grupo hidroxila do carbono anomérico de um monossacarídeo e um grupo nucleofílico — geralmente outra hidroxila — de uma segunda molécula. Esse processo envolve a eliminação de uma molécula de água e a formação de uma ligação estável entre os dois açúcares. Dependendo da natureza do grupo envolvido, podem formar-se ligações O-glicosídicas (as mais comuns, envolvendo oxigênio) ou N-glicosídicas (quando o grupo nucleofílico é uma amina). [Figura]
Um aspecto crítico dessa ligação é sua configuração estereoquímica, determinada pela orientação do grupo hidroxila no carbono anomérico no momento da formação da ligação. Isso dá origem às formas α (alfa) e β (beta). Essa distinção não é trivial: ela define propriedades estruturais e funcionais completamente diferentes nas moléculas resultantes. Por exemplo, ligações α(1→4), como as presentes no amido e no glicogênio, conferem flexibilidade às cadeias polissacarídicas, favorecendo armazenamento e mobilização energética. Em contraste, ligações β(1→4), como na celulose, resultam em estruturas lineares rígidas, altamente resistentes e com função estrutural. [Figura]
Além da configuração α ou β, as ligações glicosídicas são caracterizadas pela posição dos carbonos envolvidos. A notação (1→4), (1→6) ou outras variantes indica quais átomos de carbono participam da ligação, influenciando diretamente a arquitetura tridimensional da molécula. Ligações (1→6), por exemplo, são responsáveis por pontos de ramificação em polissacarídeos como o glicogênio, aumentando a solubilidade e a acessibilidade enzimática dessas estruturas.
A especificidade das ligações glicosídicas é rigidamente controlada por enzimas. Diferentes enzimas catalisam a formação e a quebra dessas ligações com alta seletividade, reconhecendo não apenas o tipo de ligação, mas também a configuração espacial dos substratos. Essa precisão garante que as estruturas glicídicas sejam sintetizadas de maneira ordenada e funcional, evitando a formação de arranjos aleatórios que comprometeriam a eficiência biológica.
Do ponto de vista funcional, as ligações glicosídicas determinam propriedades fundamentais dos carboidratos, como digestibilidade, solubilidade e resistência mecânica. A incapacidade de muitos organismos de degradar ligações β(1→4), por exemplo, explica por que a celulose não é digerida por humanos, enquanto o amido é facilmente metabolizado. Esse aspecto tem implicações diretas em nutrição, fisiologia e até em estratégias agrícolas e industriais.
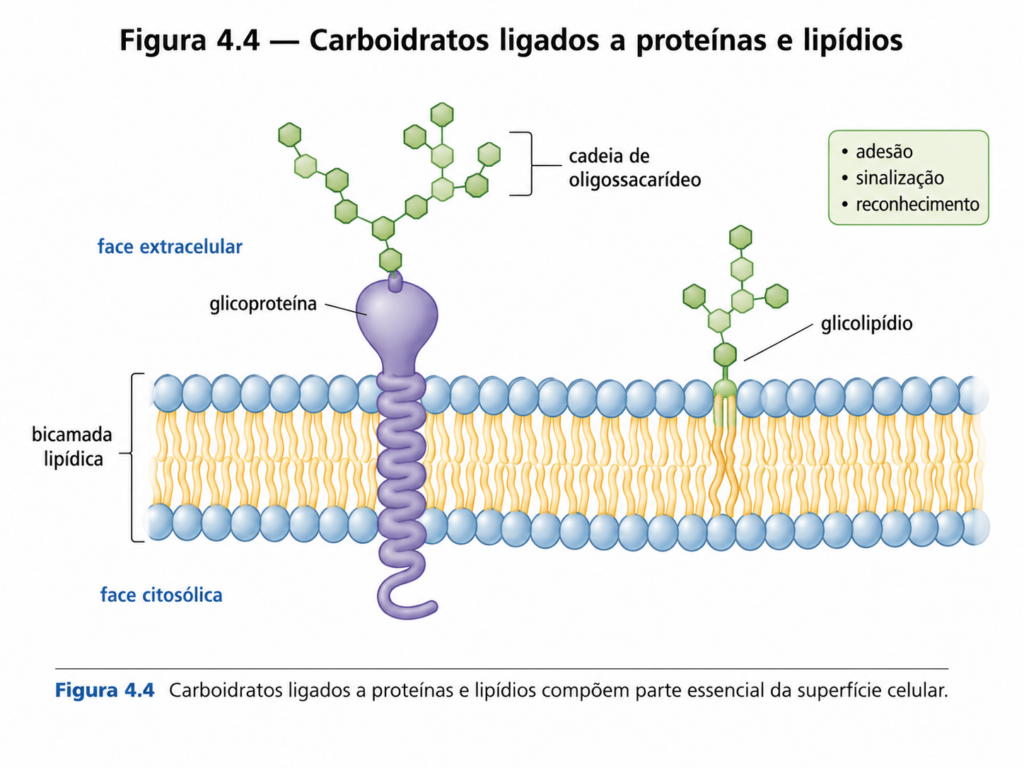
As ligações glicosídicas também desempenham papel crucial na formação de estruturas híbridas, como glicoproteínas e glicolipídios, onde carboidratos se ligam a proteínas ou lipídios. Nesses contextos, as cadeias de açúcares atuam como elementos de reconhecimento molecular, mediando interações celulares, respostas imunológicas e processos de sinalização. A precisão estrutural dessas ligações é essencial para o funcionamento adequado desses sistemas. [Figura]
Outro ponto relevante é que, ao formar uma ligação glicosídica, o carbono anomérico perde sua capacidade de interconverter entre as formas α e β, tornando-se estruturalmente fixo. Isso tem implicações diretas na reatividade química do composto, especialmente na distinção entre açúcares redutores e não redutores — conceito fundamental para entender propriedades químicas e testes laboratoriais relacionados aos carboidratos.
Portanto, as ligações glicosídicas não são apenas conexões entre unidades de açúcar; elas são elementos estruturais determinantes que definem a organização, a função e o comportamento dos carboidratos nos sistemas biológicos. Compreender sua formação, especificidade e implicações permite avançar da simples descrição molecular para uma interpretação mecanística das funções glicídicas, estabelecendo a base para o entendimento de processos metabólicos e estruturais mais complexos.
4.8 Dissacarídeos biologicamente relevantes #
A união de dois monossacarídeos por meio de uma ligação glicosídica dá origem aos dissacarídeos, uma classe de carboidratos que ocupa posição estratégica entre as formas simples e os polímeros complexos. Embora estruturalmente mais modestos que os polissacarídeos, os dissacarídeos desempenham papéis fundamentais no metabolismo energético, na nutrição e na fisiologia de diferentes organismos, funcionando frequentemente como formas intermediárias de transporte, armazenamento ou disponibilização de energia.
Entre os dissacarídeos mais relevantes, destaca-se a sacarose, formada pela união de uma molécula de glicose e uma de frutose. Essa ligação ocorre entre os carbonos anoméricos de ambos os monossacarídeos, resultando em uma ligação α(1→2)β. Essa característica estrutural tem uma consequência importante: a sacarose é um açúcar não redutor, pois nenhum dos carbonos anoméricos permanece livre para participar de reações de oxidação-redução. Do ponto de vista biológico, a sacarose é o principal carboidrato de transporte em plantas, sendo sintetizada nas folhas e distribuída para tecidos de crescimento e armazenamento por meio do floema. [Figura]
Outro dissacarídeo de grande relevância é a lactose, composta por glicose e galactose unidas por uma ligação β(1→4). Diferentemente da sacarose, a lactose possui um carbono anomérico livre, sendo classificada como açúcar redutor. Sua importância fisiológica é evidente no contexto da nutrição de mamíferos, onde atua como principal fonte de energia no leite. A digestão da lactose depende da enzima lactase, cuja atividade varia entre indivíduos e populações, estando associada a condições como intolerância à lactose quando ausente ou reduzida.
A maltose, por sua vez, é formada por duas unidades de glicose ligadas por uma ligação α(1→4). Esse dissacarídeo surge como intermediário na digestão do amido, sendo rapidamente convertido em glicose livre por ação enzimática. Sua presença ilustra como a hidrólise de polissacarídeos ocorre de maneira sequencial e controlada, liberando unidades menores que podem ser prontamente absorvidas e utilizadas pelas células.
A estrutura desses dissacarídeos evidencia um princípio recorrente na bioquímica dos carboidratos: a natureza da ligação glicosídica determina propriedades químicas e biológicas específicas. A diferença entre uma ligação α ou β, ou entre diferentes posições de ligação, altera significativamente a digestibilidade, a reatividade e o papel metabólico da molécula. Essa especificidade estrutural é reconhecida por enzimas altamente seletivas, que catalisam a hidrólise dessas ligações de forma precisa.
Do ponto de vista metabólico, os dissacarídeos não são utilizados diretamente pelas células em sua forma original. Antes disso, precisam ser hidrolisados em seus monossacarídeos constituintes por enzimas específicas no trato digestivo ou no interior celular. Esse processo garante que apenas unidades simples, como glicose, possam ser absorvidas e integradas às vias metabólicas centrais, como a glicólise. [Equação]
Além de sua função energética, os dissacarídeos também possuem relevância tecnológica e industrial. A sacarose, por exemplo, é amplamente utilizada como adoçante e matéria-prima em processos fermentativos, enquanto a lactose tem aplicações na indústria farmacêutica e alimentícia. Essas aplicações refletem propriedades físico-químicas específicas, como solubilidade, estabilidade e reatividade, que derivam diretamente de sua estrutura molecular.
Outro aspecto relevante é a capacidade de alguns dissacarídeos participarem de reações químicas não enzimáticas, como a reação de Maillard, que ocorre entre açúcares redutores e aminoácidos. Esse processo é fundamental na formação de sabores e aromas em alimentos, mas também possui implicações biológicas, como a glicação de proteínas em condições patológicas, particularmente no diabetes.
Portanto, os dissacarídeos representam mais do que simples combinações de monossacarídeos. Eles constituem unidades funcionais intermediárias que conectam a química estrutural dos carboidratos à sua aplicação biológica e tecnológica. A análise de suas estruturas, ligações e propriedades fornece uma base sólida para compreender tanto a digestão e o metabolismo quanto a organização de carboidratos mais complexos nos sistemas vivos.
4.9 Polissacarídeos estruturais #
À medida que a complexidade dos carboidratos aumenta, sua função biológica se expande para além do metabolismo energético, alcançando papéis estruturais essenciais na organização dos sistemas vivos. Os polissacarídeos estruturais representam uma classe de macromoléculas cuja principal função não é armazenar energia, mas fornecer suporte mecânico, proteção e integridade celular. Esses polímeros são projetados, do ponto de vista molecular, para maximizar resistência, estabilidade e durabilidade — propriedades diretamente derivadas de sua organização química e conformacional.
O exemplo mais emblemático dessa classe é a celulose, o principal componente da parede celular das plantas. Trata-se de um polímero linear formado por unidades de glicose unidas por ligações β(1→4). Essa configuração estereoquímica específica impõe uma orientação alternada das unidades monoméricas, resultando em cadeias estendidas e rigidamente alinhadas. A consequência dessa organização é a formação de extensas redes de ligações de hidrogênio entre cadeias adjacentes, que se agrupam em microfibrilas altamente resistentes. [Figura]
Essa arquitetura confere à celulose propriedades mecânicas excepcionais, permitindo que plantas sustentem sua estrutura contra forças gravitacionais e ambientais. Ao mesmo tempo, essa mesma rigidez torna a celulose altamente resistente à degradação enzimática. A maioria dos organismos, incluindo humanos, não possui enzimas capazes de hidrolisar ligações β(1→4), o que explica sua baixa digestibilidade. Em sistemas agrícolas, essa característica tem implicações diretas na nutrição animal, sendo contornada por microrganismos simbióticos capazes de degradar esse polímero.
Outro polissacarídeo estrutural de grande relevância é a quitina, presente no exoesqueleto de artrópodes e na parede celular de fungos. Estruturalmente semelhante à celulose, a quitina é composta por unidades de N-acetilglicosamina unidas também por ligações β(1→4). A presença do grupo acetilamino confere propriedades adicionais à molécula, aumentando sua rigidez e resistência. Assim como a celulose, a quitina forma estruturas fibrosas organizadas que proporcionam proteção mecânica e suporte estrutural. [Figura]
Nos tecidos animais, embora polissacarídeos estruturais não formem estruturas rígidas comparáveis à celulose ou quitina, eles desempenham funções igualmente importantes na matriz extracelular. Nesse contexto, destacam-se os glicosaminoglicanos, polímeros altamente hidratados compostos por unidades repetitivas de açúcares modificados. Essas moléculas, frequentemente associadas a proteínas formando proteoglicanos, conferem propriedades viscoelásticas aos tecidos, permitindo absorção de impacto, lubrificação e manutenção da integridade estrutural em ambientes biológicos dinâmicos.
A funcionalidade desses polissacarídeos está diretamente relacionada à sua organização tridimensional e à natureza das ligações glicosídicas. Diferentemente dos polissacarídeos de reserva, que apresentam estruturas ramificadas e acessíveis, os polissacarídeos estruturais são predominantemente lineares e altamente organizados. Essa linearidade favorece o empacotamento ordenado das cadeias, maximizando interações intermoleculares e aumentando a resistência mecânica.
Do ponto de vista bioquímico, essa distinção reflete um princípio fundamental: a função de uma macromolécula é uma consequência direta de sua estrutura. No caso dos polissacarídeos estruturais, a escolha evolutiva por ligações β e arranjos lineares não é arbitrária, mas uma solução otimizada para a construção de materiais biológicos resistentes e duráveis.
Além de sua importância biológica, esses polímeros possuem ampla relevância tecnológica e industrial. A celulose, por exemplo, é a base da indústria de papel e fibras têxteis, enquanto derivados de quitina têm aplicações em biotecnologia, medicina e agricultura. Essas aplicações exploram propriedades como biocompatibilidade, resistência e capacidade de modificação química.
Portanto, os polissacarídeos estruturais representam um exemplo claro de como a natureza utiliza princípios químicos simples para construir materiais com propriedades complexas e altamente especializadas. Ao compreender sua organização molecular, torna-se possível não apenas interpretar sua função nos sistemas biológicos, mas também explorar seu potencial em aplicações tecnológicas e produtivas, especialmente em contextos como o agronegócio e a bioindústria.
4.10 Polissacarídeos de reserva #
Enquanto os polissacarídeos estruturais são organizados para conferir resistência e estabilidade, os polissacarídeos de reserva são projetados com uma lógica bioquímica oposta: maximizar a armazenagem eficiente de energia e permitir sua mobilização rápida e controlada. Essa dualidade revela um princípio central da bioquímica: a estrutura molecular é moldada pela função que a molécula precisa desempenhar no sistema biológico.
Os dois principais representantes dessa classe são o amido, em plantas, e o glicogênio, em animais e microrganismos. Ambos são polímeros de glicose, mas diferem significativamente em sua organização estrutural e, consequentemente, em suas propriedades funcionais.
O amido é o principal carboidrato de reserva vegetal e encontra-se armazenado em organelas especializadas, como os amiloplastos. Estruturalmente, é composto por duas frações distintas: amilose e amilopectina. A amilose é um polímero linear de glicose unido por ligações α(1→4), que tende a formar estruturas helicoidais compactas. Já a amilopectina apresenta uma organização ramificada, com cadeias principais ligadas por α(1→4) e pontos de ramificação em α(1→6). Essa estrutura híbrida permite um equilíbrio entre compactação e acessibilidade enzimática. [Figura]
A organização do amido reflete a estratégia das plantas: armazenar energia de forma relativamente estável, mas ainda acessível quando necessário, como durante a germinação ou em períodos de baixa disponibilidade energética. A proporção entre amilose e amilopectina varia entre espécies vegetais, influenciando propriedades como digestibilidade e aplicação industrial, especialmente em alimentos e biotecnologia.
O glicogênio, por sua vez, é o principal polissacarídeo de reserva em animais e apresenta uma estrutura ainda mais altamente ramificada do que a amilopectina. As cadeias de glicose são unidas por ligações α(1→4), com ramificações α(1→6) ocorrendo em intervalos mais curtos. Essa arquitetura gera uma molécula extremamente compacta e altamente acessível às enzimas, permitindo a liberação rápida de glicose em resposta às demandas metabólicas. [Figura]
Essa característica é essencial para organismos que necessitam de respostas rápidas a variações energéticas, como durante exercício físico ou em situações de estresse metabólico. No fígado, o glicogênio atua como reserva para manutenção da glicemia, enquanto nos músculos fornece energia local para contração. Essa diferenciação funcional reforça o papel estratégico desse polímero na homeostase energética.
Do ponto de vista bioquímico, a presença de múltiplos pontos de ramificação no glicogênio tem uma implicação direta: aumenta o número de extremidades não redutoras disponíveis para ação enzimática. Isso permite que várias enzimas atuem simultaneamente na degradação da molécula, acelerando significativamente a liberação de glicose. Esse mecanismo ilustra uma otimização estrutural orientada para eficiência metabólica.
Comparativamente, a diferença entre amido e glicogênio não reside apenas na complexidade estrutural, mas na velocidade de mobilização energética. O glicogênio é mais dinâmico e responsivo, enquanto o amido é mais estável e adequado a estratégias de armazenamento de longo prazo. Essa distinção reflete adaptações evolutivas às necessidades energéticas específicas de plantas e animais.
Além de sua função biológica, os polissacarídeos de reserva possuem ampla relevância aplicada. O amido é uma das principais matérias-primas da indústria alimentícia e de biocombustíveis, enquanto o entendimento do metabolismo do glicogênio é fundamental na fisiologia humana e no manejo de condições metabólicas, como diabetes e doenças de armazenamento de glicogênio.
Outro aspecto importante é que esses polímeros não existem isoladamente; estão integrados a redes metabólicas complexas. Sua síntese e degradação são rigidamente reguladas por enzimas e sinais hormonais, garantindo que o armazenamento e a liberação de energia ocorram de maneira coordenada com as necessidades do organismo. [Equação]
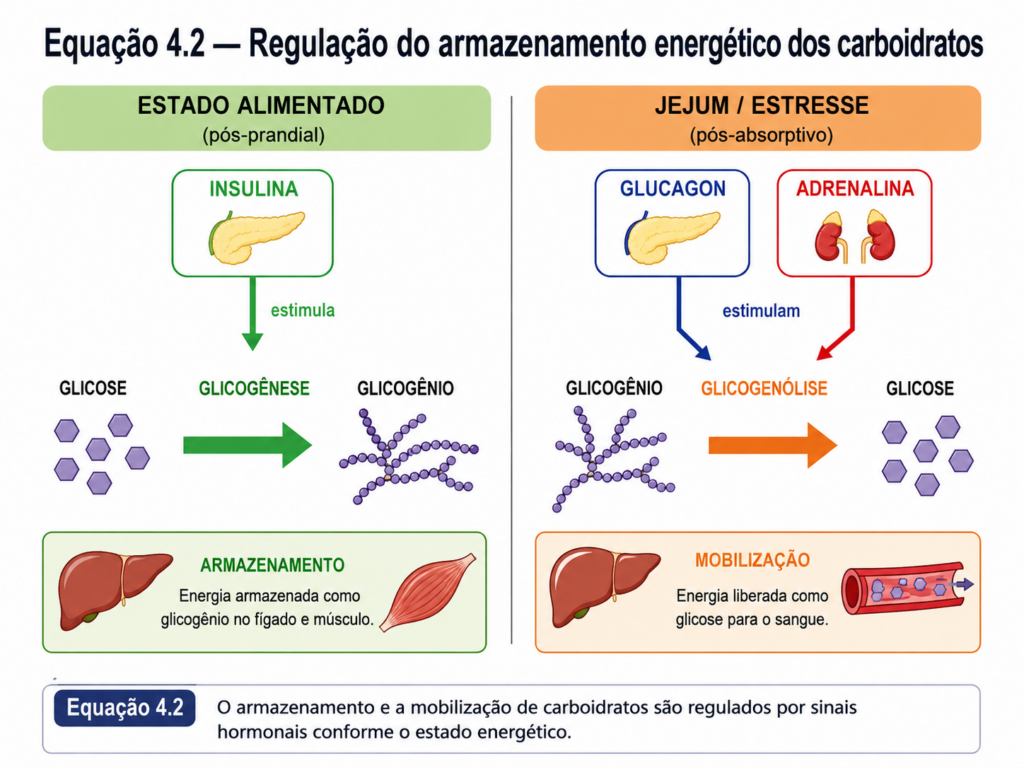
Portanto, os polissacarídeos de reserva representam um exemplo claro de como sistemas biológicos organizam moléculas para resolver um problema central: armazenar energia de forma eficiente sem comprometer a capacidade de utilizá-la rapidamente. A análise de sua estrutura e função fornece uma base essencial para compreender o metabolismo energético e sua regulação em diferentes contextos biológicos.
4.11 Propriedades físico-químicas dos carboidratos #
As propriedades físico-químicas dos carboidratos constituem a base para compreender seu comportamento em sistemas biológicos e tecnológicos. Essas propriedades emergem diretamente da combinação entre sua estrutura molecular — rica em grupos hidroxila e, em muitos casos, grupos carbonila — e sua organização espacial. Como resultado, os carboidratos apresentam características que os tornam altamente compatíveis com ambientes aquosos, quimicamente reativos e funcionalmente versáteis.
Uma das propriedades mais evidentes dos carboidratos é sua alta solubilidade em água. A presença de múltiplos grupos hidroxila permite a formação de extensas redes de ligações de hidrogênio com moléculas de água, favorecendo sua dissolução. Essa característica é essencial para o transporte e distribuição dessas moléculas no meio intracelular e extracelular. Em monossacarídeos e dissacarídeos, essa solubilidade é particularmente elevada, enquanto em polissacarídeos ela depende da estrutura — polímeros altamente organizados, como a celulose, são pouco solúveis devido à formação de redes intermoleculares rígidas. [Figura]
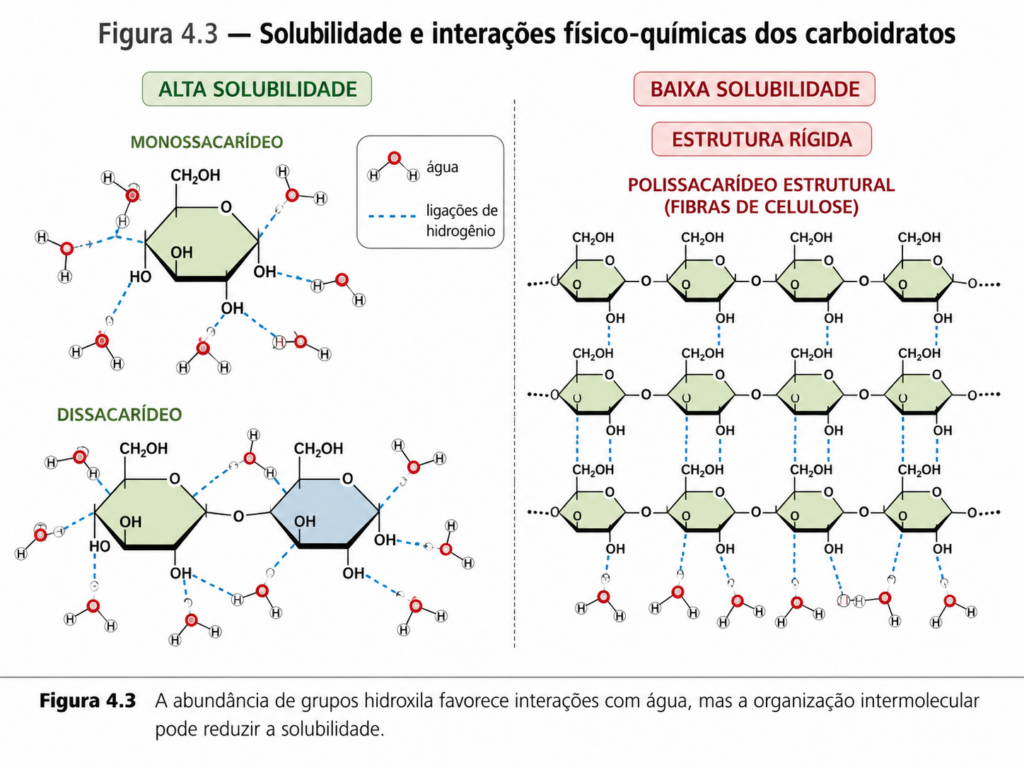
Outro aspecto central é a capacidade dos carboidratos de estabelecer interações intermoleculares. Além das ligações de hidrogênio, essas moléculas podem participar de interações dipolo-dipolo e forças de van der Waals. Em sistemas biológicos, essas interações são fundamentais para a formação de estruturas macromoleculares e para o reconhecimento molecular, especialmente em glicoproteínas e glicolipídios. Em polissacarídeos estruturais, como a celulose, essas interações resultam em materiais altamente organizados e resistentes.
Do ponto de vista químico, os carboidratos exibem significativa reatividade, particularmente devido ao grupo carbonila na forma linear e ao carbono anomérico na forma cíclica. Monossacarídeos com carbono anomérico livre são classificados como açúcares redutores, pois podem atuar como agentes redutores em reações químicas. Essa propriedade é explorada em testes laboratoriais clássicos, como as reações de Benedict e Fehling, e também tem implicações biológicas, como na glicação não enzimática de proteínas. [Equação]
As reações de oxidação e redução são especialmente relevantes. A oxidação de grupos aldeído pode gerar ácidos aldônicos, enquanto a oxidação de grupos terminais pode produzir ácidos urônicos. Já a redução do grupo carbonila resulta na formação de álcoois de açúcar, como o sorbitol. Essas transformações não apenas ampliam a diversidade estrutural dos carboidratos, mas também desempenham papéis fisiológicos importantes, incluindo processos metabólicos e mecanismos de detoxificação.
Outra propriedade importante é a capacidade de formar derivados químicos por meio de reações como esterificação e formação de ligações glicosídicas. A fosforilação de açúcares, por exemplo, altera significativamente sua reatividade e função metabólica, permitindo sua retenção no interior celular e participação em vias bioquímicas específicas. Essa modulação química evidencia a flexibilidade funcional dos carboidratos dentro das redes metabólicas.
Os carboidratos também apresentam propriedades relacionadas ao seu comportamento em solução, como a mutarrotação, que reflete o equilíbrio dinâmico entre diferentes formas anoméricas. Esse fenômeno influencia propriedades ópticas e pode afetar interações com enzimas e outras biomoléculas. Além disso, a viscosidade de soluções contendo polissacarídeos depende do tamanho e da estrutura das cadeias, sendo um fator crítico em aplicações industriais e biotecnológicas.
Do ponto de vista termodinâmico, a estabilidade dos carboidratos em solução resulta de um balanço entre interações intermoleculares e conformação estrutural. Em sistemas biológicos, essas propriedades permitem que os carboidratos atuem como moléculas dinâmicas, capazes de responder a mudanças no ambiente químico e físico.
Em aplicações práticas, essas propriedades físico-químicas são exploradas em diversas áreas. Na indústria alimentícia, influenciam textura, sabor e estabilidade de produtos. Na agricultura, afetam a digestibilidade de biomassa e a eficiência nutricional. Em biotecnologia, determinam a funcionalidade de polímeros e biomateriais.
Portanto, as propriedades físico-químicas dos carboidratos não são características isoladas, mas expressões diretas de sua estrutura molecular e organização. Compreendê-las permite interpretar o comportamento dessas moléculas em diferentes contextos, desde reações químicas simples até processos biológicos complexos, consolidando a base para o estudo integrado da bioquímica dos carboidratos.
4.12 Carboidratos na superfície celular #
À medida que a complexidade dos sistemas biológicos aumenta, os carboidratos deixam de atuar apenas como fontes de energia ou componentes estruturais e passam a desempenhar um papel mais sofisticado: mediadores de informação molecular. Na superfície celular, cadeias de carboidratos associadas a proteínas e lipídios formam uma camada altamente organizada e funcional, responsável por processos críticos como reconhecimento celular, adesão, sinalização e resposta imunológica.
Essas estruturas estão presentes principalmente na forma de glicoproteínas e glicolipídios, nos quais oligossacarídeos estão covalentemente ligados a proteínas ou lipídios da membrana plasmática. A diversidade estrutural dessas cadeias glicídicas é extraordinária, resultando de diferentes combinações de monossacarídeos, tipos de ligações glicosídicas e padrões de ramificação. Essa variabilidade confere aos carboidratos de superfície uma capacidade única de codificar informação biológica em nível molecular — um conceito frequentemente referido como “código glicídico”. [Figura]
Diferentemente do código genético, que é linear e universal, o código glicídico é tridimensional e altamente contextual. Pequenas alterações na estrutura de um oligossacarídeo — como a inversão de um grupo hidroxila ou a mudança em uma ligação glicosídica — podem alterar completamente sua capacidade de reconhecimento por proteínas específicas. Essas proteínas, conhecidas como lectinas, interagem seletivamente com determinadas estruturas glicídicas, desencadeando respostas celulares específicas.
Um dos exemplos mais claros dessa função está no sistema imunológico. Os carboidratos presentes na superfície celular atuam como marcadores de identidade, permitindo que o organismo distinga entre células próprias e estruturas estranhas. Antígenos de grupos sanguíneos, por exemplo, são determinados por variações específicas em cadeias oligossacarídicas na superfície das hemácias. Essa especificidade é crítica em processos como transfusões sanguíneas, onde incompatibilidades podem desencadear respostas imunológicas severas.
Além do reconhecimento imunológico, os carboidratos de superfície participam de processos de adesão celular. Durante o desenvolvimento embrionário, cicatrização de tecidos e formação de estruturas multicelulares, células precisam interagir de maneira coordenada. Essas interações são mediadas, em grande parte, por glicoproteínas e glicolipídios, que funcionam como pontos de ancoragem e comunicação entre células e entre células e a matriz extracelular.
Outro aspecto fundamental é o papel dos carboidratos na sinalização celular. Modificações glicídicas em proteínas de membrana podem influenciar sua estabilidade, localização e atividade funcional. Em muitos casos, a presença ou ausência de determinadas cadeias de carboidratos regula a interação dessas proteínas com seus ligantes, modulando vias de sinalização intracelular. Esse mecanismo é particularmente relevante em processos como resposta a hormônios, crescimento celular e diferenciação.
Os carboidratos também desempenham papel crítico na interação entre organismos, especialmente em contextos de infecção. Muitos patógenos reconhecem e se ligam a estruturas glicídicas específicas na superfície das células hospedeiras para iniciar processos de adesão e invasão. Em resposta, organismos evoluíram mecanismos para modificar essas estruturas, dificultando o reconhecimento por agentes infecciosos. Esse “jogo molecular” evidencia a importância dos carboidratos como interface dinâmica entre organismos e seu ambiente.
Do ponto de vista estrutural, a presença dessas cadeias glicídicas forma uma camada conhecida como glicocálix, que recobre a superfície celular. Essa camada não apenas participa de processos de reconhecimento e sinalização, mas também exerce funções físicas, como proteção mecânica e regulação da interação com o meio extracelular. [Figura]
A biossíntese dessas estruturas ocorre principalmente no retículo endoplasmático e no complexo de Golgi, onde enzimas específicas adicionam e modificam os açúcares de maneira altamente controlada. Esse processo não segue um molde direto, como na síntese de proteínas, mas depende da atividade coordenada de múltiplas enzimas, o que contribui para a diversidade estrutural observada.
Portanto, os carboidratos na superfície celular representam um nível avançado de organização bioquímica, no qual estruturas moleculares atuam como portadoras de informação e mediadoras de interação. Compreender esse sistema é essencial para interpretar fenômenos biológicos complexos, desde o funcionamento do sistema imunológico até a progressão de doenças e o desenvolvimento de estratégias terapêuticas.
4.13 Digestão, absorção e transporte de carboidratos #
A eficiência com que organismos utilizam carboidratos como fonte de energia depende de uma sequência coordenada de eventos que envolve digestão enzimática, absorção intestinal e transporte sistêmico. Esse processo transforma macromoléculas complexas em unidades simples capazes de serem incorporadas ao metabolismo celular, estabelecendo a conexão entre ingestão alimentar e produção de energia.
A digestão dos carboidratos inicia-se ainda na cavidade oral, onde a amilase salivar catalisa a hidrólise parcial das ligações α(1→4) do amido, gerando oligossacarídeos menores, como maltose e dextrinas. Embora essa etapa seja limitada pelo curto tempo de permanência do alimento na boca e pela inativação da enzima no ambiente ácido do estômago, ela representa o primeiro passo na fragmentação dessas macromoléculas.
No intestino delgado, ocorre a etapa mais significativa da digestão. A amilase pancreática, secretada no lúmen intestinal, continua a degradação do amido, produzindo dissacarídeos e oligossacarídeos. Em seguida, enzimas específicas localizadas na borda em escova das células epiteliais intestinais — como maltase, lactase e sacarase — hidrolisam esses compostos em monossacarídeos, principalmente glicose, galactose e frutose. Essa etapa final é altamente específica e essencial para que as moléculas possam ser absorvidas. [Equação]
A absorção dos monossacarídeos ocorre através de mecanismos distintos, refletindo suas propriedades químicas e necessidades fisiológicas. A glicose e a galactose são transportadas para o interior das células intestinais por meio de transporte ativo secundário, mediado pelo cotransportador SGLT1, que utiliza o gradiente de sódio como fonte de energia. Já a frutose é absorvida por difusão facilitada, via transportador GLUT5, um processo independente de sódio. [Figura]
Uma vez no interior dos enterócitos, esses monossacarídeos são transferidos para a corrente sanguínea através do transportador GLUT2, localizado na membrana basolateral. A partir daí, seguem para o fígado por meio da veia porta hepática, onde são metabolizados, armazenados ou distribuídos para outros tecidos, conforme a demanda energética do organismo.
O fígado desempenha papel central nesse processo, atuando como regulador metabólico. A glicose pode ser convertida em glicogênio para armazenamento, utilizada imediatamente na produção de energia ou liberada na circulação para manutenção da glicemia. Esse controle é essencial para garantir o fornecimento contínuo de energia, especialmente para tecidos altamente dependentes de glicose, como o sistema nervoso central.
A regulação desse sistema envolve uma complexa rede de sinais hormonais. A insulina, por exemplo, promove a captação de glicose pelos tecidos e estimula seu armazenamento, enquanto o glucagon atua de forma oposta, favorecendo a liberação de glicose no sangue. Esse equilíbrio é fundamental para a homeostase energética e, quando comprometido, pode levar a distúrbios metabólicos como o diabetes mellitus.
Do ponto de vista bioquímico, a digestão e absorção de carboidratos ilustram um princípio essencial: a necessidade de transformar macromoléculas em unidades simples para que possam atravessar membranas biológicas e participar de vias metabólicas. Esse processo não é apenas mecânico, mas altamente regulado e integrado ao estado fisiológico do organismo.
Além disso, fatores como a estrutura dos carboidratos ingeridos influenciam diretamente a eficiência desse processo. Polissacarídeos com ligações β, como a celulose, não são digeridos por enzimas humanas, sendo classificados como fibras alimentares. Essas moléculas, embora não forneçam energia diretamente, desempenham papel importante na saúde intestinal e na modulação da absorção de nutrientes.
Em contextos aplicados, especialmente na agronomia e na nutrição animal, compreender esses mecanismos é essencial para otimizar dietas e melhorar a eficiência de utilização de alimentos. A digestibilidade de diferentes fontes de carboidratos impacta diretamente o desempenho produtivo e a saúde dos organismos.
Portanto, a digestão, absorção e transporte de carboidratos constituem um sistema integrado que conecta a ingestão alimentar ao metabolismo celular. A compreensão detalhada desses processos fornece a base para interpretar tanto a fisiologia normal quanto condições patológicas, além de orientar aplicações práticas em saúde, nutrição e produção agrícola.
4.14 Integração metabólica dos carboidratos #
A análise isolada das vias metabólicas dos carboidratos — como glicólise, gliconeogênese ou metabolismo do glicogênio — fornece apenas uma visão parcial do funcionamento celular. Na realidade, essas vias estão profundamente interligadas e operam de forma coordenada dentro de uma rede metabólica integrada, ajustando-se continuamente às demandas energéticas e às condições fisiológicas do organismo. Essa integração constitui um dos pilares da bioquímica moderna, permitindo compreender como a célula mantém sua homeostase energética e funcional.
No centro dessa rede está a glicose, cuja disponibilidade e utilização são rigidamente controladas. Após sua entrada na célula, a glicose pode seguir diferentes destinos metabólicos. Em condições de alta demanda energética, é rapidamente degradada pela glicólise, gerando piruvato e, subsequentemente, intermediários do ciclo do ácido cítrico e da fosforilação oxidativa, resultando na produção de ATP. Em contrapartida, quando há excesso de energia, a glicose é direcionada para armazenamento na forma de glicogênio ou convertida em ácidos graxos, conectando o metabolismo de carboidratos ao de lipídios. [Figura]
A integração metabólica torna-se particularmente evidente no fígado, órgão central na regulação do metabolismo energético. O fígado atua como um “buffer metabólico”, captando glicose após a alimentação e liberando-a durante períodos de jejum. Esse equilíbrio é alcançado por meio da alternância entre processos anabólicos, como a glicogênese, e catabólicos, como a glicogenólise e a gliconeogênese. Essa dinâmica garante a manutenção da glicemia dentro de limites estreitos, essencial para o funcionamento adequado de tecidos dependentes de glicose.
Durante o estado alimentado, caracterizado pela abundância de nutrientes, a glicose é direcionada principalmente para armazenamento e síntese. A glicólise é ativada, e seus intermediários podem ser desviados para a síntese de lipídios e outros compostos. Nesse contexto, a insulina atua como principal regulador hormonal, estimulando a captação de glicose e promovendo vias anabólicas.
Em contraste, durante o jejum ou em situações de alta demanda energética, o organismo mobiliza suas reservas. O glicogênio hepático é degradado, liberando glicose para a corrente sanguínea. À medida que essas reservas se esgotam, a gliconeogênese torna-se a principal fonte de glicose, utilizando precursores como aminoácidos, lactato e glicerol. Esse processo evidencia a interconexão entre o metabolismo de carboidratos, proteínas e lipídios. [Equação]
Outro exemplo de integração metabólica é o ciclo de Cori, no qual o lactato produzido nos músculos durante exercício anaeróbio é transportado ao fígado, onde é convertido novamente em glicose. Esse mecanismo permite a reutilização de intermediários metabólicos e a manutenção da produção de energia em diferentes tecidos, demonstrando a cooperação sistêmica entre órgãos.
A via das pentoses fosfato também desempenha papel relevante nessa integração, desviando glicose-6-fosfato para a produção de NADPH e ribose-5-fosfato. O NADPH é essencial para reações biossintéticas e para a defesa antioxidante, enquanto a ribose-5-fosfato é necessária para a síntese de nucleotídeos. Assim, os carboidratos não apenas fornecem energia, mas também participam da biossíntese de moléculas fundamentais.
A regulação dessas vias ocorre em múltiplos níveis, incluindo controle alostérico de enzimas, modificação covalente e regulação hormonal. Enzimas-chave, como a fosfofrutoquinase-1 na glicólise e a glicose-6-fosfatase na gliconeogênese, funcionam como pontos de controle que respondem a sinais energéticos e hormonais. Essa regulação garante que vias opostas não operem simultaneamente de forma ineficiente, evitando desperdício energético.
Do ponto de vista sistêmico, a integração metabólica dos carboidratos permite que o organismo responda de maneira adaptativa a diferentes condições fisiológicas, como alimentação, jejum, exercício e estresse. Essa flexibilidade metabólica é essencial para a sobrevivência, permitindo o uso eficiente de recursos energéticos disponíveis.
Em contextos patológicos, falhas nessa integração podem levar a distúrbios metabólicos significativos. O diabetes mellitus, por exemplo, resulta de alterações na regulação da glicose, comprometendo sua captação e utilização pelos tecidos. Isso evidencia que a integração metabólica não é apenas um conceito teórico, mas um componente crítico da saúde fisiológica.
Portanto, a integração metabólica dos carboidratos representa a convergência entre múltiplas vias bioquímicas, coordenadas por mecanismos regulatórios sofisticados. Compreender essa rede permite transcender a análise isolada de reações e alcançar uma visão sistêmica do metabolismo, essencial para interpretar fenômenos biológicos e desenvolver aplicações em saúde, agronomia e biotecnologia.
4.15 Aplicações dos carboidratos em saúde, agro e indústria #
A relevância dos carboidratos transcende o domínio da bioquímica fundamental e se manifesta de forma concreta em áreas estratégicas como saúde, produção agrícola e processos industriais. Essa amplitude de aplicação decorre diretamente de suas propriedades estruturais, reatividade química e papel central no metabolismo, tornando-os moléculas-chave na interface entre ciência básica e inovação tecnológica.
No campo da saúde, os carboidratos ocupam posição central na regulação metabólica e no diagnóstico de doenças. A glicose, como principal substrato energético, é rigidamente controlada no organismo, e alterações nesse controle estão associadas a patologias como o diabetes mellitus. A monitorização da glicemia e a análise de marcadores como a hemoglobina glicada são ferramentas fundamentais na prática clínica, refletindo diretamente o metabolismo dos carboidratos. Além disso, a modificação não enzimática de proteínas por açúcares redutores — processo conhecido como glicação — está relacionada ao desenvolvimento de complicações crônicas, evidenciando o impacto bioquímico dessas moléculas na fisiopatologia.
Outro aspecto relevante é o papel dos carboidratos na nutrição e na microbiota intestinal. Polissacarídeos não digeríveis, como fibras alimentares, modulam a atividade microbiana no intestino, influenciando processos metabólicos, imunológicos e até mesmo neurológicos. Esse entendimento tem impulsionado o desenvolvimento de alimentos funcionais e estratégias nutricionais voltadas à promoção da saúde.
Na agricultura, os carboidratos são determinantes na produtividade e na qualidade das culturas. A fotossíntese, responsável pela síntese de açúcares a partir de dióxido de carbono e energia luminosa, sustenta toda a produção vegetal. O acúmulo e a distribuição de carboidratos nas plantas influenciam diretamente o crescimento, o rendimento e a resistência a estresses ambientais. Em culturas agrícolas, a manipulação do metabolismo de carboidratos — seja por melhoramento genético ou manejo nutricional — pode otimizar a produção de biomassa e a eficiência no uso de recursos.
Além disso, a composição de carboidratos em alimentos vegetais impacta diretamente a nutrição animal e humana. A digestibilidade de polissacarídeos, como amido e fibras, influencia o aproveitamento energético e o desempenho produtivo, especialmente em sistemas de produção no semiárido, onde a eficiência no uso de recursos é crítica. Nesse contexto, compreender a bioquímica dos carboidratos permite desenvolver estratégias mais eficientes e sustentáveis de produção.
No setor industrial, os carboidratos constituem uma das principais matérias-primas para uma ampla gama de aplicações. O amido, por exemplo, é utilizado na produção de alimentos, bioplásticos, adesivos e biocombustíveis. Sua estrutura pode ser modificada quimicamente ou enzimaticamente para atender a diferentes demandas tecnológicas, como controle de viscosidade, estabilidade e gelatinização.
A fermentação de açúcares é outro processo de grande relevância industrial, sendo a base para a produção de etanol, ácidos orgânicos, bebidas alcoólicas e diversos bioprodutos. Nesse contexto, monossacarídeos e dissacarídeos são convertidos por microrganismos em compostos de interesse econômico, integrando bioquímica e biotecnologia em cadeias produtivas estratégicas.
Os carboidratos também têm aplicações emergentes em biotecnologia e medicina, incluindo o desenvolvimento de biomateriais, sistemas de liberação controlada de fármacos e engenharia de tecidos. Polissacarídeos como a celulose modificada e a quitina apresentam propriedades de biocompatibilidade e biodegradabilidade que os tornam altamente atrativos para essas aplicações.
Outro campo em expansão é o da glicobiologia aplicada, que explora o papel dos carboidratos em processos de reconhecimento molecular para o desenvolvimento de vacinas, terapias e diagnósticos. A especificidade das interações entre carboidratos e proteínas abre possibilidades para intervenções direcionadas em doenças infecciosas, câncer e distúrbios metabólicos.
Do ponto de vista estratégico, o domínio sobre a bioquímica dos carboidratos permite integrar conhecimento científico com aplicação prática, gerando soluções inovadoras em diferentes setores. Em regiões como o semiárido brasileiro, essa integração pode ser particularmente relevante, possibilitando o desenvolvimento de tecnologias adaptadas às condições locais e promovendo maior eficiência produtiva.
Portanto, os carboidratos não devem ser vistos apenas como componentes básicos da matéria viva, mas como recursos versáteis e estratégicos, cuja compreensão profunda permite explorar seu potencial em múltiplas dimensões. A articulação entre estrutura, função e aplicação consolida esses compostos como elementos centrais tanto na ciência quanto na inovação tecnológica.
Referências #
NELSON, David L.; COX, Michael M. Princípios de bioquímica de Lehninger. 6. ed. Porto Alegre: Artmed, 2014.
VOET, Donald; VOET, Judith G.; PRATT, Charlotte W. Fundamentos de bioquímica: a vida em nível molecular. 4. ed. Porto Alegre: Artmed, 2014.
STRYER, Lubert; BERG, Jeremy M.; TYMOCZKO, John L.; GATTO JR., Gregory J. Bioquímica. 8. ed. Rio de Janeiro: Guanabara Koogan, 2014.
MURRAY, Robert K. et al. Bioquímica ilustrada de Harper. 30. ed. Porto Alegre: AMGH, 2017.
Estudo dirigido — Capítulo 4: Carboidratos #
- Explique por que os carboidratos podem atuar tanto como fonte de energia quanto como componentes estruturais. Use como exemplos a glicose, o amido, o glicogênio e a celulose.
- Diferencie monossacarídeos, dissacarídeos, oligossacarídeos e polissacarídeos. Em sua resposta, indique pelo menos um exemplo biológico de cada grupo.
- Por que pequenas diferenças na estrutura dos carboidratos podem gerar funções biológicas muito diferentes? Relacione sua resposta com isomeria, anômeros α e β e ligações glicosídicas.
- Compare amido, glicogênio e celulose quanto à estrutura e à função. Explique por que o amido e o glicogênio são moléculas de reserva energética, enquanto a celulose é uma molécula estrutural.
- Explique como os carboidratos participam do reconhecimento celular. Relacione glicoproteínas, glicolipídios, glicocálix e especificidade molecular.
Pergunta para provocar pesquisa #
- Pesquise como a estrutura dos carboidratos influencia aplicações práticas em saúde, agricultura ou indústria. Escolha um exemplo — diabetes, fibras alimentares, bioetanol, biomateriais de celulose, quitina/quitosana, prebióticos ou produtividade vegetal — e explique como a propriedade molecular do carboidrato se conecta à aplicação escolhida.



