A função proteica depende diretamente de sua conformação tridimensional. Este capítulo aprofunda a relação entre estrutura e atividade, abordando proteínas globulares e fibrosas, além dos efeitos da desnaturação. A estabilidade estrutural é analisada como resultado de interações químicas específicas, permitindo entender como alterações ambientais impactam a função biológica.
7.1 Organização hierárquica das proteínas: da sequência à função #
A compreensão das proteínas como entidades funcionais exige, antes de tudo, o abandono de uma visão simplista que as reduz a estruturas estáticas. Proteínas são sistemas dinâmicos cuja função emerge de uma organização hierárquica altamente sofisticada, iniciada em uma sequência linear de aminoácidos e culminando em arranjos tridimensionais capazes de interagir com precisão molecular em ambientes biológicos complexos. Essa hierarquia estrutural não é apenas uma classificação didática, mas uma expressão direta dos princípios físico-químicos que governam a matéria viva.
O ponto de partida dessa organização é a sequência de aminoácidos, determinada pela informação genética codificada no DNA e traduzida em cadeias polipeptídicas. Cada aminoácido, com suas propriedades químicas específicas — polaridade, carga, tamanho e capacidade de formar ligações — contribui para um código estrutural implícito. Esse código não é explícito como o genético, mas manifesta-se na tendência da cadeia polipeptídica em adotar conformações energeticamente favoráveis. A sequência, portanto, contém toda a informação necessária para o dobramento da proteína, um princípio clássico da bioquímica estrutural que sustenta a ideia de que estrutura é função potencial em estado latente.
À medida que a cadeia polipeptídica emerge durante a síntese ribossomal, começam a se formar padrões locais de organização conhecidos como estruturas secundárias. Esses arranjos, como as α-hélices e folhas β, resultam de interações regulares entre grupos peptídicos ao longo do esqueleto da cadeia, especialmente por meio de ligações de hidrogênio. Essas estruturas não surgem ao acaso, mas refletem restrições geométricas impostas pela planaridade da ligação peptídica e pelos ângulos torsionais permitidos, constituindo os primeiros níveis de organização espacial previsível da proteína [Figura].
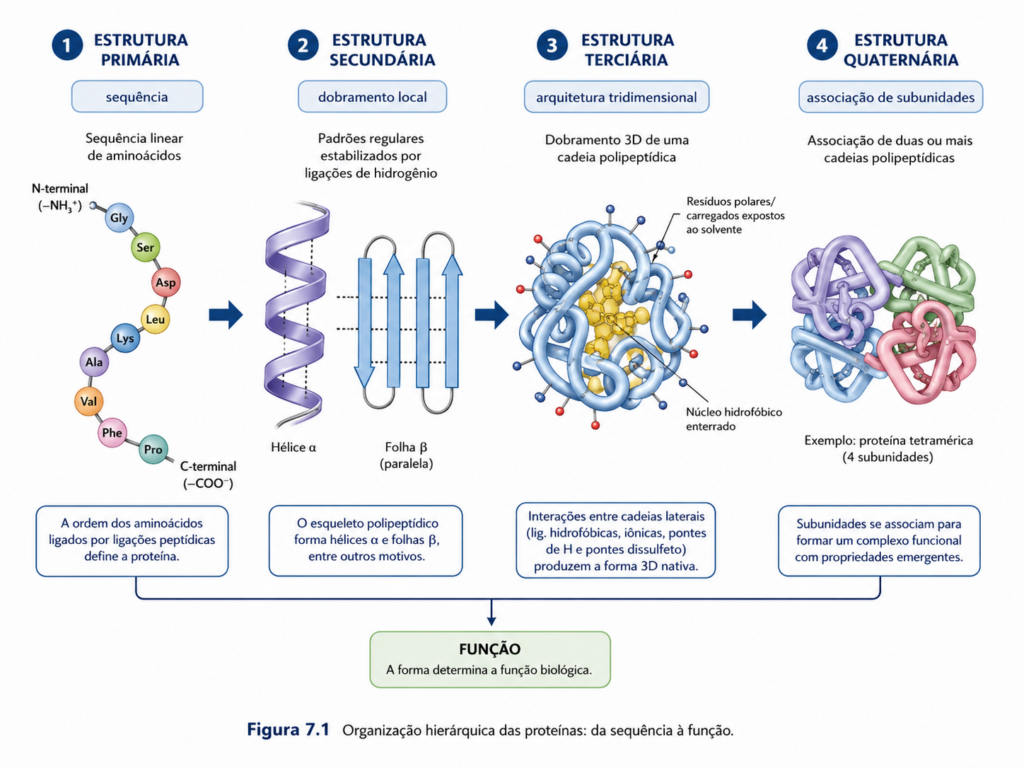
No entanto, a funcionalidade plena das proteínas só se estabelece quando essas estruturas locais se organizam em uma arquitetura tridimensional completa, denominada estrutura terciária. Nesse nível, cadeias laterais de aminoácidos interagem por meio de forças não covalentes — interações hidrofóbicas, ligações de hidrogênio, interações iônicas e forças de van der Waals — além de eventuais ligações covalentes, como pontes dissulfeto. O ambiente aquoso desempenha papel central nesse processo, favorecendo o colapso hidrofóbico que impulsiona o dobramento da proteína em direção a um estado de mínima energia livre. Assim, a estrutura terciária representa a materialização física da informação contida na sequência primária, transformando potencial em função efetiva.
Em muitas proteínas, especialmente aquelas envolvidas em processos regulatórios ou estruturais complexos, a organização não se limita a uma única cadeia polipeptídica. Surge então a estrutura quaternária, caracterizada pela associação de múltiplas subunidades. Essa organização permite propriedades emergentes que não seriam possíveis em proteínas monoméricas, como a cooperatividade e o controle alostérico. A interação entre subunidades cria sistemas moleculares capazes de responder de forma sensível a variações no ambiente celular, ajustando sua atividade conforme a necessidade fisiológica [Figura].
O aspecto mais relevante dessa organização hierárquica é que ela não é linear no sentido funcional. Embora descrita em níveis — primário, secundário, terciário e quaternário —, esses elementos operam de forma integrada e interdependente. Alterações mínimas na sequência podem provocar mudanças significativas na estrutura tridimensional, afetando diretamente a função da proteína. Esse princípio explica desde mutações patológicas até a adaptação evolutiva de proteínas a diferentes condições ambientais.
Além disso, a organização estrutural das proteínas deve ser compreendida como um estado dinâmico, sujeito a flutuações conformacionais contínuas. Mesmo em sua forma nativa, proteínas não são rígidas; elas oscilam entre microestados estruturais que possibilitam interações com ligantes, substratos e outras macromoléculas. Essa flexibilidade é essencial para a atividade biológica e estabelece a base para fenômenos como reconhecimento molecular e catálise enzimática.
Portanto, a organização hierárquica das proteínas constitui o elo fundamental entre informação genética e função biológica. Ela traduz uma sequência linear de aminoácidos em máquinas moleculares altamente especializadas, capazes de executar tarefas com precisão e eficiência notáveis. Compreender essa hierarquia não é apenas descrever níveis estruturais, mas entender como a vida, em nível molecular, emerge da interação ordenada entre estrutura, energia e dinâmica [Quadro].
7.2 Estrutura primária e implicações funcionais #
A estrutura primária de uma proteína corresponde à sequência linear de aminoácidos unidos por ligações peptídicas, formando uma cadeia polipeptídica com direção definida, do terminal amino (N-terminal) ao terminal carboxila (C-terminal). À primeira vista, essa definição pode sugerir um nível estrutural simples e desprovido de complexidade funcional. No entanto, essa interpretação é equivocada. A estrutura primária representa o nível mais fundamental de organização molecular das proteínas, pois contém, de forma codificada, todas as informações necessárias para o dobramento tridimensional e, consequentemente, para a atividade biológica.
Cada aminoácido inserido na cadeia contribui com propriedades físico-químicas específicas que influenciam diretamente o comportamento da proteína. A natureza das cadeias laterais — hidrofóbicas, polares, carregadas positiva ou negativamente — determina padrões de interação que orientarão o processo de dobramento. Resíduos hidrofóbicos tendem a se agrupar no interior da proteína, afastando-se do meio aquoso, enquanto resíduos polares e carregados permanecem, em geral, expostos ao solvente. Essa distribuição não é aleatória, mas guiada por princípios termodinâmicos que favorecem estados de menor energia livre [Figura].
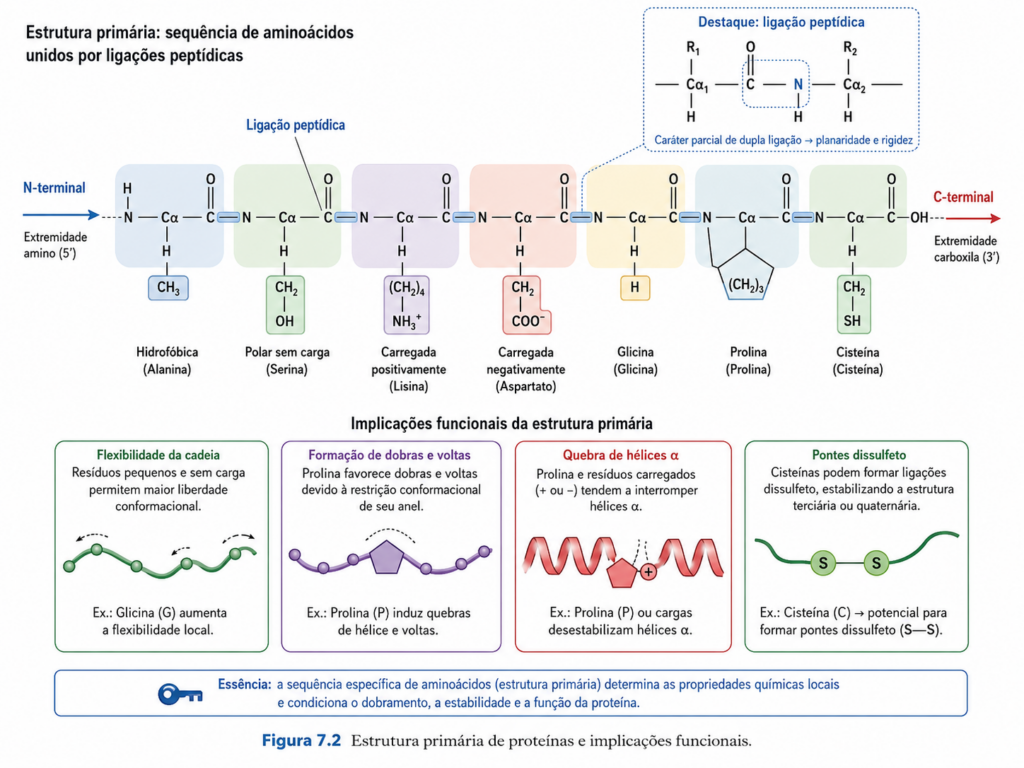
A sequência primária também impõe restrições estruturais importantes. Certos aminoácidos desempenham papéis estruturais específicos. A prolina, por exemplo, devido à sua estrutura cíclica, introduz descontinuidades e quebras em hélices α, funcionando como um elemento disruptivo na regularidade estrutural. Já a glicina, por possuir uma cadeia lateral mínima, confere elevada flexibilidade à cadeia polipeptídica, sendo frequentemente encontrada em regiões de dobra ou em sítios onde a mobilidade estrutural é essencial. A cisteína, por sua vez, é capaz de formar pontes dissulfeto, estabelecendo ligações covalentes que estabilizam a estrutura tridimensional, especialmente em proteínas extracelulares.
Além das propriedades individuais dos aminoácidos, a ordem em que esses resíduos aparecem na sequência é determinante. Pequenas alterações, como substituições pontuais, podem provocar efeitos significativos na estrutura e na função da proteína. Um exemplo clássico é a substituição de um único aminoácido na cadeia β da hemoglobina, que resulta na formação da hemoglobina S, associada à anemia falciforme. Nesse caso, a troca de um resíduo polar por um hidrofóbico altera as interações intermoleculares, levando à agregação anormal das proteínas e à deformação das hemácias. Esse fenômeno ilustra de forma inequívoca como a estrutura primária influencia diretamente propriedades macroscópicas e fisiológicas [Quadro].
A estrutura primária também reflete a história evolutiva das proteínas. Sequências conservadas entre diferentes organismos indicam regiões críticas para a função, frequentemente associadas a sítios ativos ou regiões de ligação a ligantes. Por outro lado, regiões mais variáveis podem estar relacionadas a adaptações específicas ou funções regulatórias. A comparação de sequências proteicas tornou-se, assim, uma ferramenta central para inferir relações evolutivas, prever funções e identificar domínios estruturais conservados.
Outro aspecto relevante é que a estrutura primária não é apenas determinada geneticamente, mas pode sofrer modificações pós-traducionais. Processos como fosforilação, glicosilação, acetilação e clivagem proteolítica alteram quimicamente determinados resíduos, expandindo a diversidade funcional das proteínas sem necessidade de alteração na sequência codificada no DNA. Essas modificações introduzem um nível adicional de complexidade, permitindo que uma mesma proteína desempenhe múltiplas funções dependendo do contexto celular.
Do ponto de vista termodinâmico, a estrutura primária define o “espaço de conformações” acessível à proteína. Embora o dobramento seja um processo complexo, envolvendo múltiplos intermediários, ele não ocorre de forma aleatória. A sequência direciona a proteína para um estado nativo específico, geralmente correspondente a um mínimo global ou quase global de energia livre. Esse princípio, amplamente demonstrado em estudos clássicos de dobramento proteico, reforça a ideia de que a informação estrutural está intrinsecamente codificada na sequência de aminoácidos.
Portanto, a estrutura primária deve ser compreendida como o determinante central da identidade molecular de uma proteína. Ela não apenas define a composição química da cadeia polipeptídica, mas estabelece as bases para todos os níveis subsequentes de organização estrutural e funcional. Alterações nesse nível repercutem em cascata sobre toda a arquitetura da proteína, afetando sua estabilidade, dinâmica e capacidade de interação. Em última instância, a estrutura primária conecta o código genético à função biológica, constituindo o primeiro elo na cadeia que transforma informação molecular em atividade vital [Figura].
7.3 Estruturas secundárias: α-hélice, folhas β e elementos estruturais recorrentes #
A transição entre a linearidade da estrutura primária e a complexidade tridimensional das proteínas inicia-se com a formação de padrões locais altamente organizados conhecidos como estruturas secundárias. Esses arranjos representam soluções geométricas e energéticas para a organização do esqueleto polipeptídico, emergindo de interações regulares e repetitivas entre os grupos amida (–NH) e carbonila (–C=O) da cadeia principal. Diferentemente das interações envolvendo cadeias laterais, a estrutura secundária é governada principalmente pela própria arquitetura da ligação peptídica, cuja planaridade e rigidez impõem restrições conformacionais específicas.
A ligação peptídica apresenta caráter parcial de dupla ligação, resultado da ressonância entre o carbono carbonílico e o nitrogênio amídico. Essa característica limita a rotação ao redor dessa ligação, tornando o plano peptídico essencialmente rígido. Assim, a flexibilidade da cadeia polipeptídica reside nos ângulos torsionais φ (phi) e ψ (psi), que definem a orientação relativa entre os planos peptídicos consecutivos. Nem todas as combinações desses ângulos são energeticamente viáveis; apenas determinadas regiões do espaço conformacional são permitidas, como evidenciado pelo diagrama de Ramachandran [Figura]. É dentro dessas regiões que emergem os padrões estruturais mais estáveis: a α-hélice e a folha β.
A α-hélice constitui um dos elementos estruturais mais comuns e estáveis nas proteínas. Nesse arranjo, a cadeia polipeptídica assume uma conformação helicoidal destrogira, na qual cada grupo carbonila forma uma ligação de hidrogênio com o grupo amida localizado quatro resíduos à frente na sequência (i → i+4). Essa organização gera uma estrutura compacta, com 3,6 resíduos por volta e um passo de aproximadamente 5,4 Å. As cadeias laterais projetam-se para fora da hélice, minimizando interferências estéricas e permitindo interações com o ambiente ou com outras partes da proteína. A estabilidade da α-hélice depende da natureza dos aminoácidos presentes; resíduos como alanina favorecem sua formação, enquanto prolina, devido à sua rigidez e incapacidade de formar ligações de hidrogênio adequadas, atua como um agente desestabilizador, frequentemente interrompendo a hélice [Figura].
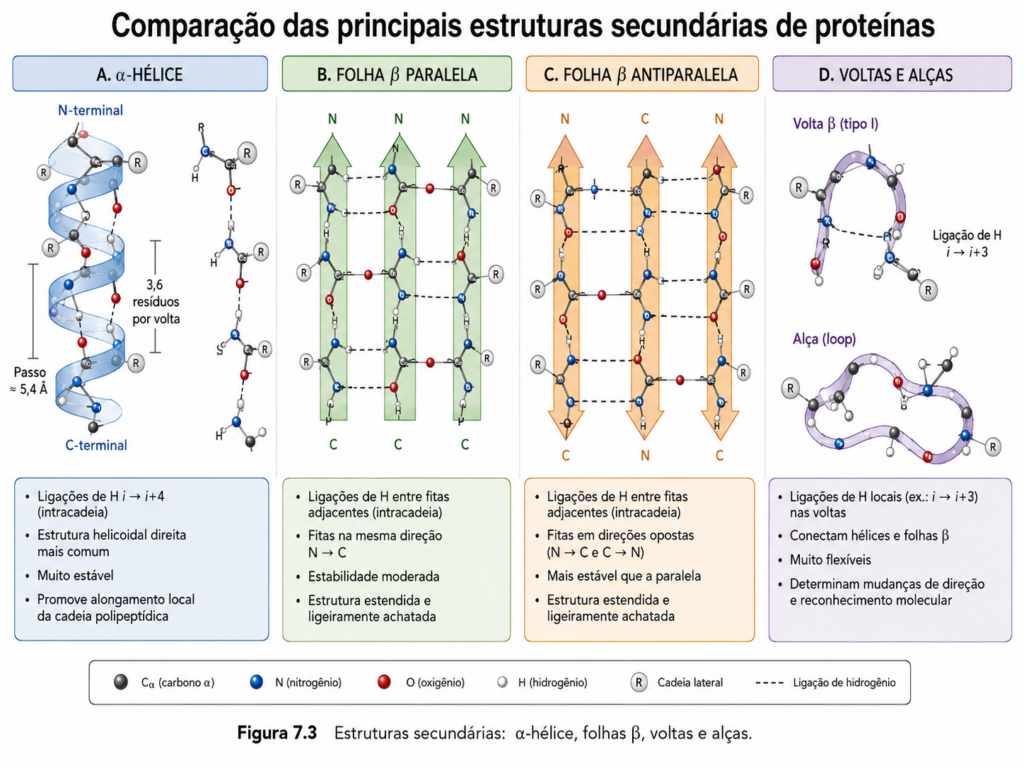
As folhas β representam um segundo padrão fundamental de organização secundária, caracterizado por cadeias polipeptídicas estendidas que se alinham lado a lado, formando uma rede de ligações de hidrogênio entre segmentos adjacentes. Essas cadeias podem estar orientadas no mesmo sentido (paralelas) ou em sentidos opostos (antiparalelas), sendo esta última configuração geralmente mais estável devido à geometria mais favorável das ligações de hidrogênio. A estrutura resultante não é completamente plana, apresentando uma leve torção que confere estabilidade adicional ao arranjo. As cadeias laterais alternam-se acima e abaixo do plano da folha, permitindo uma organização eficiente das interações com o meio e com outras regiões da proteína [Figura].
Além desses elementos clássicos, existem estruturas secundárias menos regulares, mas igualmente importantes para a arquitetura proteica. As voltas (turns) e alças (loops) conectam segmentos de α-hélice e folhas β, permitindo mudanças de direção na cadeia polipeptídica. Essas regiões são frequentemente localizadas na superfície das proteínas e desempenham papéis críticos na interação com outras moléculas, incluindo substratos, ligantes e outras proteínas. A presença de glicina e prolina é comum nessas regiões, devido à sua capacidade de conferir flexibilidade ou induzir curvaturas acentuadas na cadeia.
A estrutura secundária não deve ser interpretada como um estágio isolado do dobramento proteico, mas como parte de um processo cooperativo e integrado. Elementos secundários frequentemente se organizam em motivos estruturais recorrentes, como hélice-volta-hélice, folhas β em forma de barril (β-barrel) ou motivos β-α-β, que servem como blocos de construção para estruturas mais complexas. Esses motivos são conservados ao longo da evolução e estão associados a funções específicas, como ligação ao DNA, formação de poros em membranas ou reconhecimento molecular.
Do ponto de vista termodinâmico, a formação de estruturas secundárias representa um compromisso entre a maximização de interações favoráveis (ligações de hidrogênio) e a minimização de repulsões estéricas. Embora essas interações sejam individualmente fracas, sua repetição ao longo da cadeia confere estabilidade significativa ao conjunto. No entanto, essa estabilidade é modulada pelo contexto estrutural mais amplo, incluindo interações terciárias e condições ambientais, como pH, temperatura e composição do solvente.
Assim, as estruturas secundárias constituem o primeiro nível de organização tridimensional efetiva das proteínas, estabelecendo padrões espaciais que orientam o dobramento global. Elas funcionam como unidades estruturais intermediárias, conectando a informação contida na sequência primária à complexidade funcional da estrutura terciária. Compreender esses padrões não é apenas descrever formas geométricas, mas reconhecer os princípios físico-químicos que permitem à matéria biológica organizar-se de maneira precisa e funcional [Quadro].
7.4 Estrutura terciária: dobramento proteico e domínios funcionais #
A estrutura terciária representa o nível em que a proteína adquire sua identidade tridimensional completa, transformando uma cadeia polipeptídica linear em uma entidade funcional capaz de reconhecer, interagir e catalisar processos biológicos com elevada especificidade. Nesse estágio, os elementos de estrutura secundária — α-hélices, folhas β, voltas e alças — organizam-se em um arranjo espacial único, determinado pela soma das interações entre cadeias laterais e pelo contexto físico-químico do meio celular. É nesse nível que a proteína deixa de ser apenas uma sequência organizada e passa a atuar como uma máquina molecular.
O dobramento proteico é um processo altamente coordenado, guiado por princípios termodinâmicos que favorecem a formação de um estado de menor energia livre. Esse estado, conhecido como conformação nativa, resulta de um delicado equilíbrio entre forças estabilizadoras e desestabilizadoras. Entre as interações mais relevantes estão o efeito hidrofóbico, que promove o agrupamento de resíduos apolares no interior da proteína; as ligações de hidrogênio, que estabilizam arranjos específicos; as interações iônicas entre grupos carregados; e as forças de van der Waals, que contribuem para o empacotamento eficiente dos átomos. Em alguns casos, ligações covalentes, como pontes dissulfeto entre resíduos de cisteína, adicionam estabilidade adicional, especialmente em proteínas expostas a ambientes extracelulares [Figura].
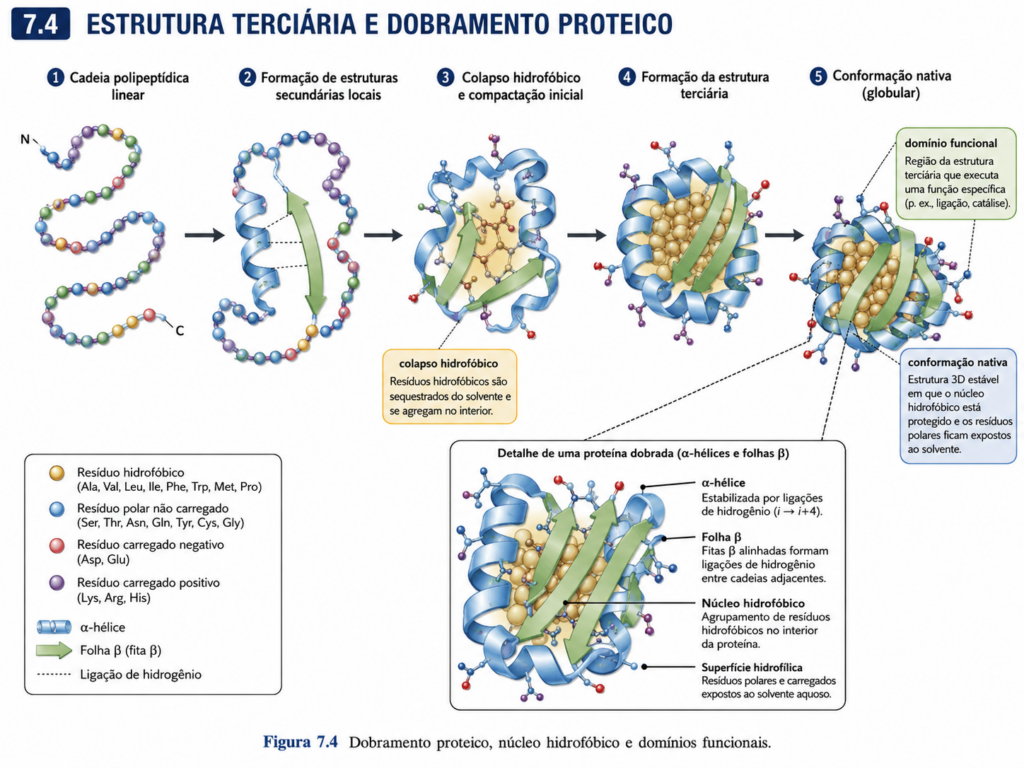
Um aspecto central da estrutura terciária é a formação de um núcleo hidrofóbico compacto. Em meio aquoso, resíduos apolares tendem a se afastar da água, levando ao colapso da cadeia polipeptídica em torno de um interior pouco acessível ao solvente. Esse colapso não é aleatório; ele é orientado pela sequência de aminoácidos e pelas restrições conformacionais impostas pelas estruturas secundárias previamente formadas. O resultado é uma arquitetura tridimensional altamente específica, na qual resíduos hidrofóbicos permanecem protegidos no interior, enquanto resíduos polares e carregados se distribuem preferencialmente na superfície, permitindo interação com o meio e com outras moléculas.
Dentro dessa organização global, emergem unidades estruturais e funcionais conhecidas como domínios. Um domínio é uma região da proteína que pode dobrar-se de forma relativamente independente, apresentando uma estrutura estável e frequentemente associada a uma função específica. Proteínas podem conter um ou vários domínios, e a combinação desses módulos permite a construção de sistemas moleculares complexos a partir de unidades relativamente simples. Domínios de ligação a DNA, domínios catalíticos e domínios de interação proteína-proteína são exemplos recorrentes na biologia molecular. Essa modularidade facilita a evolução de novas funções, uma vez que domínios podem ser recombinados, duplicados ou modificados ao longo do tempo evolutivo [Figura].
A relação entre estrutura terciária e função é direta e inseparável. O sítio ativo de uma enzima, por exemplo, não é definido por uma sequência linear de aminoácidos, mas por sua disposição tridimensional no espaço. Resíduos que estão distantes na sequência primária podem aproximar-se no espaço tridimensional para formar um ambiente químico altamente especializado, capaz de estabilizar estados de transição e acelerar reações químicas. Esse princípio também se aplica a proteínas de ligação, como receptores e anticorpos, nos quais a complementaridade estrutural entre proteína e ligante determina a especificidade da interação.
Apesar de sua aparente estabilidade, a estrutura terciária não é rígida. Proteínas apresentam flutuações conformacionais contínuas, explorando um conjunto de microestados estruturais próximos da conformação nativa. Essa dinâmica é essencial para a função, permitindo que proteínas ajustem sua forma em resposta à ligação de substratos ou a mudanças no ambiente. Fenômenos como o modelo de ajuste induzido (induced fit) ilustram como a estrutura terciária pode se reorganizar para otimizar interações moleculares, aumentando a eficiência catalítica ou a afinidade por ligantes [Quadro].
O processo de dobramento proteico, embora espontâneo em muitos casos, pode ser assistido por proteínas especializadas conhecidas como chaperonas moleculares. Essas moléculas não determinam a estrutura final da proteína, mas evitam interações incorretas durante o dobramento, especialmente em ambientes celulares densamente povoados por macromoléculas. A falha nesse processo pode levar à formação de agregados proteicos insolúveis, associados a diversas patologias, como doenças neurodegenerativas. Isso evidencia que a estrutura terciária não é apenas uma questão de estabilidade física, mas também de controle biológico rigoroso.
Do ponto de vista energético, o dobramento de proteínas ilustra um dos problemas clássicos da bioquímica: como uma cadeia polipeptídica encontra rapidamente sua conformação nativa entre um número astronomicamente grande de possibilidades conformacionais. A solução para esse paradoxo reside na existência de caminhos de dobramento preferenciais, nos quais intermediários estruturais conduzem a proteína de forma eficiente ao estado final. Esses caminhos refletem a paisagem energética da proteína, caracterizada por um funil de energia que direciona o sistema para estados cada vez mais estáveis [Figura].
Portanto, a estrutura terciária constitui o ponto de convergência entre sequência, energia e função. Ela traduz a informação linear codificada no DNA em uma arquitetura tridimensional capaz de executar tarefas específicas com alta precisão. Compreender esse nível de organização é essencial para interpretar mecanismos enzimáticos, processos de reconhecimento molecular e, em última instância, a lógica funcional dos sistemas biológicos.
7.5 Estrutura quaternária e cooperação entre subunidades #
A estrutura quaternária representa o nível de organização em que múltiplas cadeias polipeptídicas — denominadas subunidades — se associam para formar um complexo proteico funcional. Diferentemente da estrutura terciária, que descreve o arranjo tridimensional de uma única cadeia, a estrutura quaternária emerge da interação coordenada entre duas ou mais subunidades, podendo ser idênticas (homoméricas) ou distintas (heteroméricas). Essa organização não é meramente estrutural; ela introduz propriedades funcionais que não existem em proteínas monoméricas, configurando um dos exemplos mais claros de comportamento emergente na bioquímica.
A associação entre subunidades é estabilizada predominantemente por interações não covalentes — hidrofóbicas, eletrostáticas, ligações de hidrogênio e forças de van der Waals — semelhantes às que estabilizam a estrutura terciária. Em alguns casos, ligações covalentes, como pontes dissulfeto intercadeias, também podem estar presentes, especialmente em proteínas extracelulares. A interface entre subunidades é altamente específica, resultando em complementaridade geométrica e química que permite o reconhecimento seletivo entre as partes do complexo [Figura].
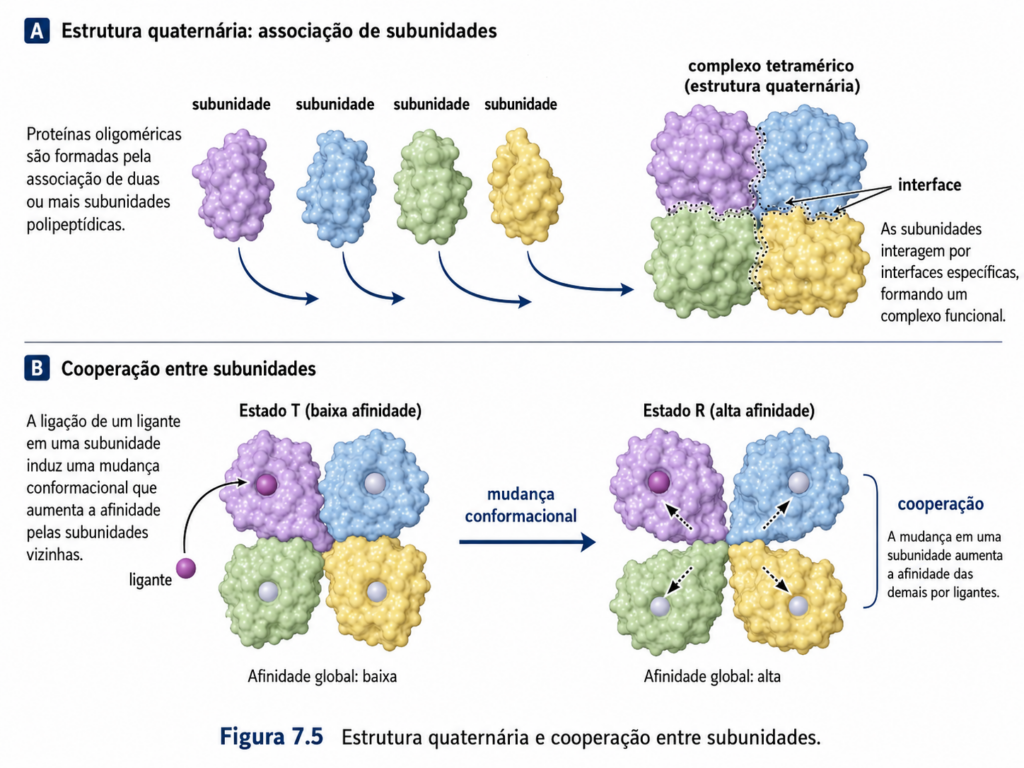
Um aspecto central da estrutura quaternária é a simetria. Muitos complexos proteicos apresentam arranjos simétricos, como dímeros, trímeros ou tetrâmeros organizados em eixos de rotação definidos. Essa simetria não apenas facilita a montagem do complexo, reduzindo a necessidade de codificação genética de múltiplas cadeias diferentes, mas também contribui para a estabilidade estrutural e para a eficiência funcional. A repetição de subunidades idênticas é uma estratégia evolutiva eficiente para construir sistemas moleculares complexos com economia de informação genética.
A principal vantagem funcional da estrutura quaternária reside na possibilidade de cooperação entre subunidades. Esse fenômeno ocorre quando a interação de uma subunidade com um ligante influencia a afinidade das demais subunidades pelo mesmo ligante. O exemplo clássico é a hemoglobina, uma proteína tetramérica responsável pelo transporte de oxigênio no sangue. A ligação de uma molécula de O₂ a uma subunidade induz mudanças conformacionais que aumentam a afinidade das outras subunidades pelo oxigênio, caracterizando a chamada cooperatividade positiva. Esse comportamento resulta em uma curva sigmoide de saturação, permitindo uma resposta altamente eficiente às variações de concentração de oxigênio no organismo [Figura].
Esse tipo de comunicação entre subunidades está frequentemente associado ao alosterismo, no qual a ligação de um ligante em um sítio específico (sítio alostérico) provoca alterações estruturais que modulam a atividade da proteína em outro sítio, geralmente o sítio ativo. A estrutura quaternária fornece o arcabouço necessário para esse tipo de regulação, permitindo que sinais moleculares sejam transmitidos através da proteína por meio de mudanças conformacionais coordenadas. Assim, proteínas oligoméricas atuam como sensores e reguladores dinâmicos de processos celulares.
Outro aspecto relevante é que a dissociação e associação de subunidades podem ser, em si, mecanismos regulatórios. Em determinadas condições fisiológicas, proteínas podem alternar entre formas oligoméricas e monoméricas, modulando sua atividade de acordo com o contexto celular. Esse comportamento adiciona uma camada adicional de controle, permitindo respostas rápidas a mudanças ambientais sem necessidade de síntese ou degradação proteica.
Do ponto de vista estrutural, a montagem de complexos quaternários requer precisão na orientação espacial das subunidades. Pequenas alterações na interface de interação podem comprometer a estabilidade do complexo ou alterar sua função. Mutações que afetam essas interfaces podem levar a disfunções significativas, evidenciando a importância da integridade estrutural nesse nível de organização. Em sistemas biológicos, essa precisão é garantida por processos de dobramento e montagem altamente regulados, frequentemente assistidos por chaperonas moleculares.
Além disso, a estrutura quaternária amplia significativamente a diversidade funcional das proteínas. Complexos multiproteicos podem integrar diferentes atividades em uma única unidade funcional, como ocorre em máquinas moleculares complexas — ribossomos, proteassomos e complexos enzimáticos multissubunidades. Nesses sistemas, a proximidade espacial entre subunidades facilita a transferência de intermediários metabólicos, aumentando a eficiência e reduzindo perdas por difusão [Quadro].
Do ponto de vista energético, a formação de estruturas quaternárias reflete um balanço entre forças de associação e entropia. Embora a associação de subunidades reduza a entropia do sistema, essa perda é compensada por interações favoráveis que estabilizam o complexo. Em muitos casos, a montagem é reversível, permitindo que a proteína responda dinamicamente às condições do meio.
Portanto, a estrutura quaternária não deve ser vista apenas como um nível adicional de organização, mas como um mecanismo estratégico para amplificação funcional. Ela permite que proteínas operem de forma cooperativa, regulada e integrada, expandindo significativamente o repertório funcional dos sistemas biológicos. A partir desse nível, a proteína deixa de ser uma unidade isolada e passa a atuar como parte de um sistema coletivo, no qual a interação entre subunidades define o comportamento global da molécula.
7.6 Forças estabilizadoras das proteínas: interações não covalentes e covalentes #
A estabilidade estrutural das proteínas não é resultado de uma única força dominante, mas da integração de múltiplas interações químicas que, individualmente, são relativamente fracas, porém, em conjunto, conferem elevada robustez à conformação nativa. Esse equilíbrio dinâmico entre forças estabilizadoras e desestabilizadoras define não apenas a integridade estrutural da proteína, mas também sua capacidade de responder a variações ambientais sem perda funcional. Compreender essas interações é essencial para interpretar como proteínas se dobram, mantêm sua forma e, ao mesmo tempo, preservam a flexibilidade necessária para sua atividade biológica.
Entre as forças mais determinantes está o efeito hidrofóbico, frequentemente considerado o principal motor do dobramento proteico em meio aquoso. Resíduos apolares tendem a se agrupar no interior da proteína, afastando-se da água, o que reduz a organização das moléculas de solvente ao redor desses grupos e, consequentemente, aumenta a entropia do sistema. Esse colapso hidrofóbico leva à formação de um núcleo interno compacto, que constitui a espinha dorsal da estrutura terciária. Mais do que uma interação direta entre grupos hidrofóbicos, trata-se de um fenômeno emergente da reorganização do solvente, sendo, portanto, fundamentalmente entrópico [Figura].
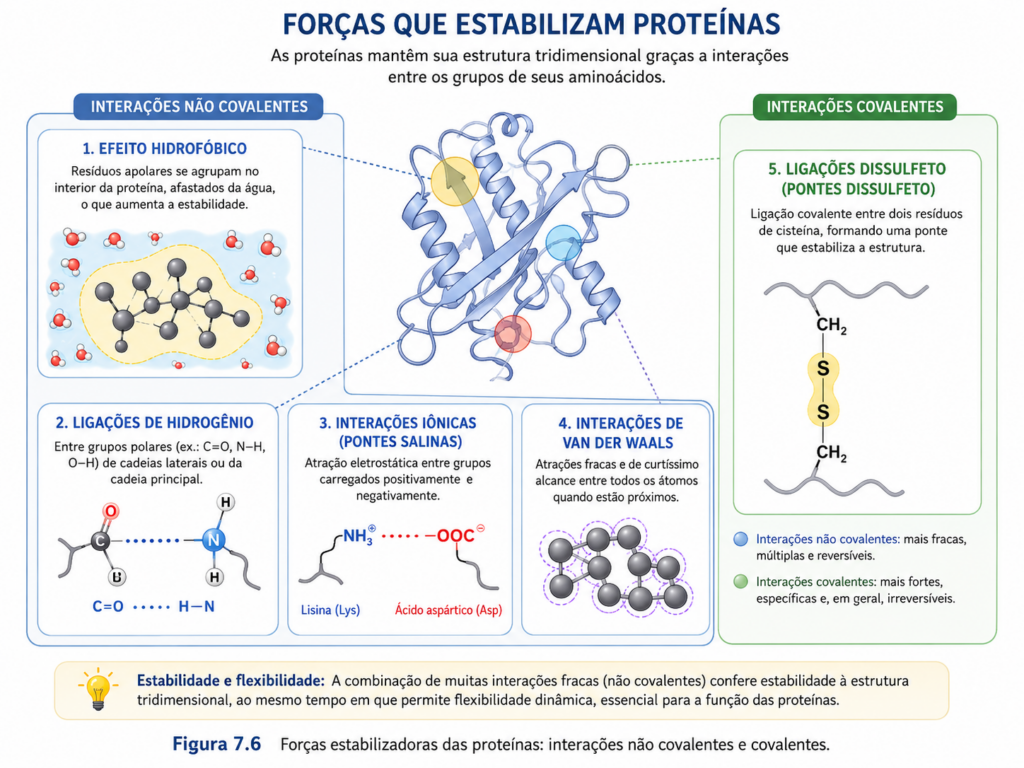
As ligações de hidrogênio desempenham papel complementar importante, especialmente na estabilização de estruturas secundárias como α-hélices e folhas β. Essas interações ocorrem entre átomos eletronegativos, como oxigênio e nitrogênio, e hidrogênios parcialmente positivos, contribuindo para a orientação precisa da cadeia polipeptídica. Embora relativamente fracas em comparação com ligações covalentes, sua abundância e organização repetitiva conferem estabilidade significativa ao conjunto estrutural. No interior da proteína, onde o ambiente é menos polar, essas ligações tornam-se ainda mais relevantes para a manutenção da conformação [Figura].
As interações eletrostáticas, ou ligações iônicas, ocorrem entre grupos carregados positivamente (como lisina e arginina) e negativamente (como aspartato e glutamato). Essas interações podem formar pares iônicos ou pontes salinas, contribuindo para a estabilização local da estrutura. Sua intensidade depende fortemente do pH e da força iônica do meio, tornando-as particularmente sensíveis a alterações ambientais. Em superfícies proteicas, essas interações também participam de processos de reconhecimento molecular e associação entre proteínas.
As forças de van der Waals, embora individualmente muito fracas, tornam-se relevantes quando há empacotamento eficiente de átomos no interior da proteína. Essas interações dependem da proximidade entre átomos e contribuem para a complementaridade estérica da estrutura tridimensional. O ajuste fino entre cadeias laterais no núcleo proteico maximiza essas interações, favorecendo estados energeticamente mais estáveis. Pequenas imperfeições nesse empacotamento podem resultar em cavidades ou tensões estruturais que comprometem a estabilidade global.
Além das interações não covalentes, algumas proteínas apresentam ligações covalentes que reforçam sua estrutura. As mais importantes são as pontes dissulfeto, formadas pela oxidação de dois resíduos de cisteína. Essas ligações são particularmente comuns em proteínas secretadas ou localizadas em ambientes extracelulares, onde as condições oxidantes favorecem sua formação. As pontes dissulfeto atuam como “travamentos estruturais”, restringindo a mobilidade da cadeia e aumentando a resistência da proteína a condições adversas, como variações de temperatura e pH [Figura].
Outro tipo de estabilização covalente pode ocorrer por meio de modificações pós-traducionais, como ligações cruzadas entre resíduos ou associação com grupos prostéticos. Essas modificações expandem a diversidade estrutural e funcional das proteínas, permitindo adaptações específicas a diferentes contextos biológicos.
A estabilidade proteica deve ser entendida como um estado termodinâmico relativo, no qual a conformação nativa é favorecida, mas não absoluta. Proteínas estão em constante equilíbrio com estados parcialmente dobrados ou desenovelados, e esse equilíbrio pode ser deslocado por fatores externos, como temperatura, pH, solventes e presença de agentes desnaturantes. A desnaturação ocorre quando as interações estabilizadoras são perturbadas, levando à perda da estrutura tridimensional e, consequentemente, da função biológica. Em alguns casos, esse processo é reversível, permitindo que a proteína retorne ao estado nativo, desde que as condições adequadas sejam restabelecidas [Quadro].
É importante destacar que a estabilidade não deve ser confundida com rigidez. Proteínas funcionalmente eficientes apresentam um equilíbrio entre estabilidade estrutural e flexibilidade dinâmica. Interações muito fortes poderiam comprometer a capacidade da proteína de sofrer mudanças conformacionais necessárias para sua atividade, enquanto interações muito fracas levariam à instabilidade estrutural. A natureza encontrou, ao longo da evolução, um ponto ótimo nesse balanço, permitindo que proteínas sejam simultaneamente estáveis e responsivas.
Portanto, as forças estabilizadoras das proteínas constituem a base físico-química que sustenta toda a arquitetura molecular dessas biomoléculas. Elas integram efeitos entrópicos e entálpicos, interações locais e globais, e contribuem para a formação de estruturas altamente organizadas e funcionalmente competentes. Compreender essas forças é essencial não apenas para descrever a estrutura proteica, mas para prever seu comportamento, manipular sua função e interpretar os efeitos de alterações genéticas ou ambientais sobre sistemas biológicos complexos.
7.7 Dinâmica conformacional e plasticidade estrutural #
A ideia de que proteínas possuem uma única estrutura fixa e rigidamente definida é incompatível com a realidade molecular observada em sistemas biológicos. Mesmo em seu estado nativo, proteínas existem como ensembles dinâmicos, explorando continuamente um conjunto de microestados conformacionais próximos em energia. Essa dinâmica não é um ruído estrutural irrelevante; ela constitui um elemento central da função proteica. A atividade biológica de uma proteína depende, em grande medida, da sua capacidade de alternar entre diferentes conformações de maneira controlada e responsiva ao ambiente.
Do ponto de vista termodinâmico, a estrutura de uma proteína deve ser entendida como um mínimo em uma paisagem energética complexa, frequentemente descrita como um “funil de energia”. Nesse modelo, múltiplas conformações parcialmente dobradas convergem para um conjunto de estados nativos energeticamente favoráveis, mas não necessariamente únicos. A existência de estados quase equivalentes permite flutuações estruturais contínuas, impulsionadas pela energia térmica do sistema. Essas flutuações ocorrem em diferentes escalas de tempo — desde movimentos rápidos de cadeias laterais até rearranjos mais lentos de domínios inteiros [Figura].
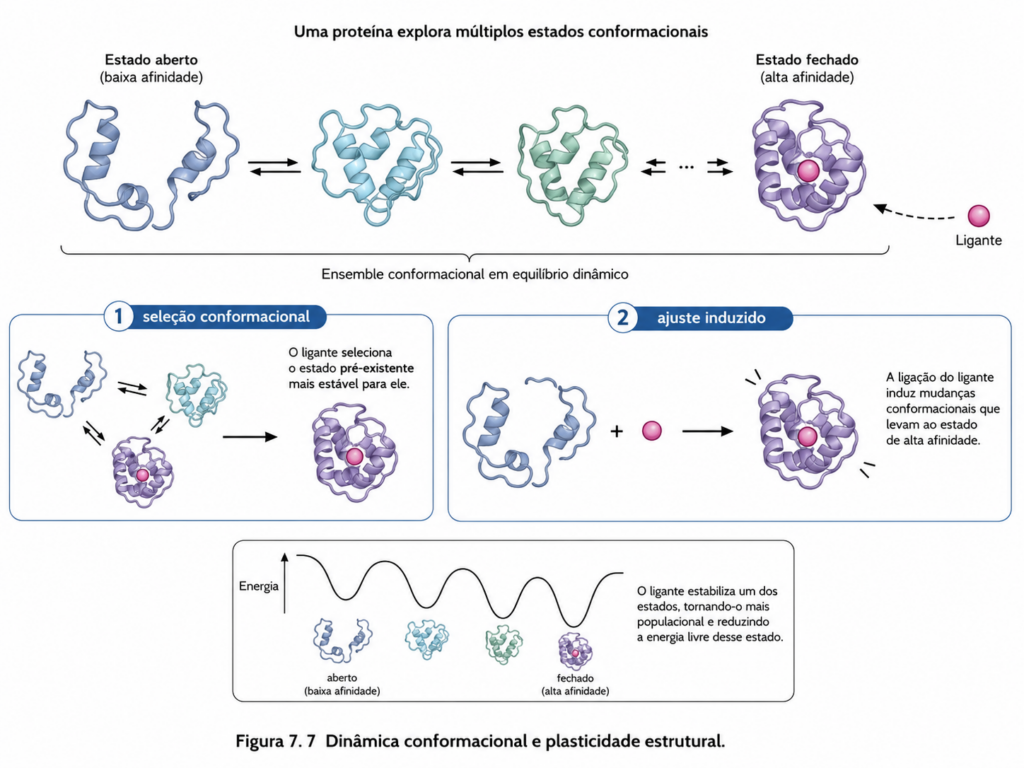
A plasticidade estrutural manifesta-se de forma particularmente evidente em proteínas que interagem com ligantes. O reconhecimento molecular não é um processo puramente estático, baseado apenas na complementaridade estrutural pré-existente. Em muitos casos, a ligação de um ligante induz mudanças conformacionais na proteína, ajustando sua estrutura para otimizar a interação — um fenômeno descrito pelo modelo de ajuste induzido. Alternativamente, no modelo de seleção conformacional, a proteína já existe em múltiplas conformações, e o ligante estabiliza aquela mais favorável à ligação. Na prática, esses dois mecanismos não são mutuamente exclusivos, mas representam extremos de um espectro de comportamentos dinâmicos [Quadro].
Essa capacidade de reorganização estrutural é fundamental para a catálise enzimática. Durante uma reação, a enzima não apenas fornece um sítio ativo adequado, mas frequentemente passa por mudanças conformacionais que estabilizam estados de transição, posicionam substratos e facilitam a liberação de produtos. A dinâmica conformacional permite que diferentes etapas do ciclo catalítico sejam coordenadas de forma eficiente, transformando a enzima em um sistema molecular adaptativo, e não em uma estrutura passiva.
A plasticidade também é essencial em processos regulatórios. Proteínas alostéricas, por exemplo, dependem de mudanças conformacionais para transmitir sinais entre diferentes regiões da molécula. A ligação de um efetor em um sítio específico pode induzir rearranjos estruturais que modulam a atividade em outro sítio distante. Esse tipo de comunicação intramolecular só é possível porque a proteína possui flexibilidade suficiente para propagar alterações estruturais ao longo de sua arquitetura tridimensional.
Em um nível mais amplo, a dinâmica conformacional está associada à existência de regiões estruturalmente menos organizadas, conhecidas como regiões intrinsecamente desordenadas. Diferentemente das estruturas clássicas bem definidas, essas regiões não adotam uma conformação única estável, permanecendo flexíveis e adaptáveis. Essa característica permite interações com múltiplos parceiros e participação em processos regulatórios complexos, especialmente em redes de sinalização celular. Longe de representar uma imperfeição estrutural, o desordenamento controlado é uma estratégia funcional altamente eficiente.
A influência do ambiente sobre a dinâmica proteica é igualmente relevante. Fatores como temperatura, pH, força iônica e presença de ligantes modulam o equilíbrio entre diferentes estados conformacionais. Pequenas variações nessas condições podem deslocar esse equilíbrio, alterando a atividade da proteína. Esse comportamento confere sensibilidade às proteínas, permitindo que atuem como sensores moleculares capazes de responder a mudanças no meio celular.
Do ponto de vista experimental, a dinâmica conformacional pode ser investigada por técnicas como ressonância magnética nuclear, espectroscopia de fluorescência, cristalografia de raios X em múltiplos estados e microscopia crioeletrônica. Essas abordagens revelam que proteínas frequentemente existem em múltiplas conformações mesmo em condições aparentemente estáveis, reforçando a ideia de que a função biológica está intrinsecamente ligada à flexibilidade estrutural.
A perda ou alteração da dinâmica conformacional pode ter consequências graves. Proteínas excessivamente rígidas podem perder eficiência funcional, enquanto proteínas excessivamente flexíveis podem tornar-se instáveis ou propensas à agregação. Em diversas doenças, como distúrbios neurodegenerativos, a desregulação da dinâmica proteica leva à formação de agregados insolúveis, evidenciando a importância do equilíbrio entre estabilidade e flexibilidade.
Portanto, a dinâmica conformacional e a plasticidade estrutural devem ser vistas como propriedades essenciais das proteínas, e não como exceções. Elas permitem que proteínas adaptem sua forma às exigências funcionais, integrem sinais moleculares e realizem processos complexos com precisão. Em última análise, é essa capacidade de movimento controlado que transforma estruturas moleculares em sistemas vivos, conectando forma, energia e função em uma única unidade operacional [Figura].
7.8 Proteínas globulares versus fibrosas: estrutura e papel biológico #
A diversidade funcional das proteínas está intimamente associada às diferenças em sua organização estrutural global. Entre os padrões mais fundamentais dessa diversidade destacam-se dois grandes grupos: as proteínas globulares e as proteínas fibrosas. Essa distinção não é meramente morfológica, mas reflete estratégias estruturais distintas que atendem a demandas biológicas específicas. Enquanto as proteínas globulares são tipicamente associadas a funções dinâmicas e metabólicas, as proteínas fibrosas desempenham papéis estruturais e mecânicos, conferindo resistência e integridade aos tecidos biológicos.
As proteínas globulares caracterizam-se por uma conformação compacta e aproximadamente esférica, resultante do dobramento tridimensional que leva à formação de um núcleo hidrofóbico bem definido e uma superfície predominantemente hidrofílica. Essa organização favorece a solubilidade em meio aquoso, condição essencial para proteínas que atuam no citosol, no plasma ou em compartimentos celulares internos. A arquitetura globular permite a formação de sítios ativos altamente específicos, nos quais resíduos de aminoácidos são posicionados com precisão para interagir com substratos, ligantes ou outras macromoléculas [Figura].
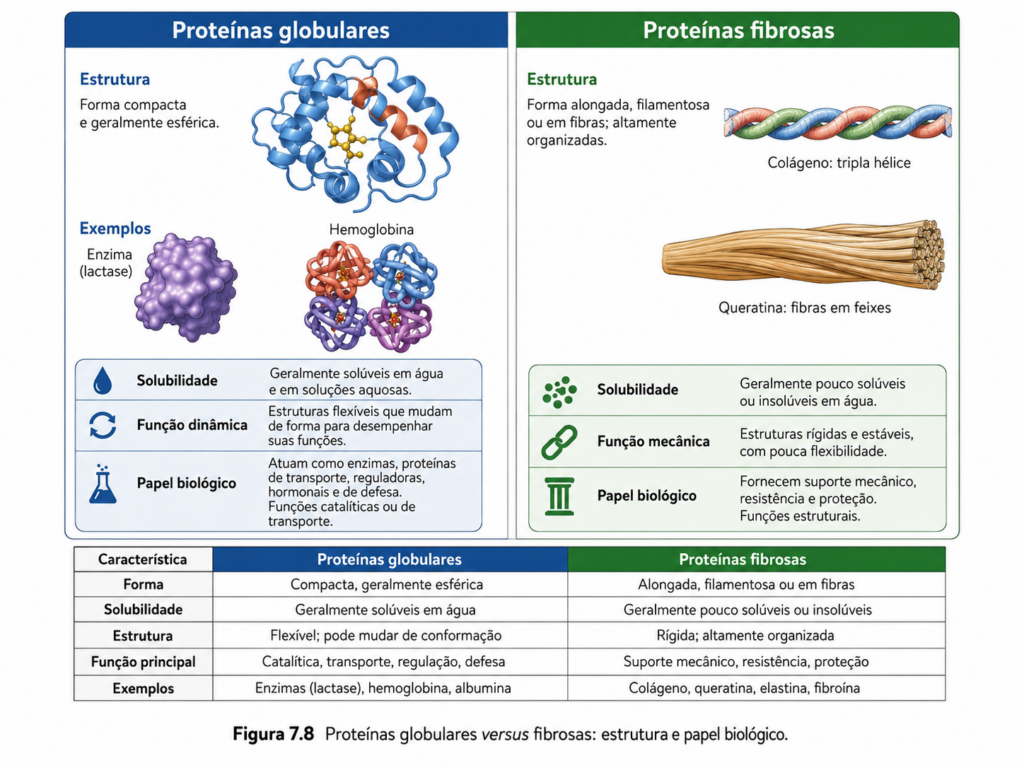
Essa classe inclui enzimas, proteínas de transporte, anticorpos e diversas proteínas regulatórias. A funcionalidade dessas proteínas depende diretamente de sua capacidade de sofrer mudanças conformacionais, o que reforça a importância da dinâmica estrutural discutida anteriormente. A flexibilidade relativa das proteínas globulares permite ajustes finos na interação com ligantes, viabilizando mecanismos como catálise enzimática, reconhecimento molecular e regulação alostérica. Assim, sua estrutura compacta não implica rigidez, mas sim uma organização eficiente que equilibra estabilidade e mobilidade.
Em contraste, as proteínas fibrosas apresentam uma organização alongada e repetitiva, frequentemente baseada em padrões regulares de estrutura secundária. Essas proteínas são, em geral, insolúveis em água e formam agregados estruturais estáveis que desempenham funções mecânicas. Diferentemente das globulares, as fibrosas não possuem um núcleo hidrofóbico bem definido nem uma conformação tridimensional compacta; sua estrutura é dominada por repetições que se estendem ao longo de grandes distâncias, formando fibras ou redes estruturais [Figura].
Um exemplo clássico é o colágeno, a proteína mais abundante em organismos animais, responsável pela resistência mecânica de tecidos como pele, ossos e tendões. Sua estrutura baseia-se em uma tripla hélice formada por três cadeias polipeptídicas entrelaçadas, estabilizadas por ligações de hidrogênio e interações específicas entre resíduos de glicina, prolina e hidroxiprolina. Outro exemplo é a queratina, presente em cabelos, unhas e epiderme, cuja estrutura helicoidal é reforçada por pontes dissulfeto, conferindo alta resistência e elasticidade. Já a fibroína da seda apresenta folhas β altamente organizadas, responsáveis por sua resistência à tração.
A diferença funcional entre proteínas globulares e fibrosas decorre diretamente de suas propriedades estruturais. Proteínas globulares são adaptadas para interações específicas e transitórias, essenciais em processos metabólicos e regulatórios. Já as proteínas fibrosas são projetadas para suportar tensões mecânicas e manter a integridade estrutural de células e tecidos. Essa especialização estrutural reflete uma otimização evolutiva, na qual diferentes arranjos moleculares são selecionados de acordo com a função desempenhada.
Apesar dessa distinção clara, é importante reconhecer que a fronteira entre esses dois grupos não é absolutamente rígida. Algumas proteínas apresentam características intermediárias ou combinam domínios globulares e regiões fibrosas em uma mesma molécula. Além disso, proteínas globulares podem formar agregados estruturais sob determinadas condições, enquanto proteínas fibrosas podem apresentar regiões com maior flexibilidade funcional. Essa plasticidade reforça a ideia de que a classificação é útil do ponto de vista didático, mas não deve ser interpretada como uma dicotomia absoluta.
Do ponto de vista aplicado, a compreensão dessas diferenças é particularmente relevante. Em sistemas agrícolas, por exemplo, proteínas fibrosas estão diretamente associadas à resistência estrutural de tecidos vegetais, influenciando características como textura e durabilidade. Já proteínas globulares, como enzimas, desempenham papéis centrais em processos metabólicos que impactam produtividade, crescimento e resposta a estresses ambientais. Na indústria, essas propriedades são exploradas em materiais biomiméticos, alimentos e biotecnologia.
Portanto, a distinção entre proteínas globulares e fibrosas evidencia como a forma molecular é moldada pela função. Enquanto as globulares operam como máquinas químicas altamente especializadas, as fibrosas atuam como componentes estruturais essenciais à organização dos sistemas biológicos. Ambas representam soluções estruturais eficientes para desafios distintos, demonstrando a capacidade da bioquímica de explorar princípios físico-químicos para gerar diversidade funcional a partir de um conjunto limitado de componentes moleculares [Quadro].
7.9 Relação estrutura–função em proteínas: princípios gerais #
A relação entre estrutura e função constitui o princípio organizador central da bioquímica das proteínas. Não se trata de uma correlação superficial, mas de uma dependência causal direta: a função de uma proteína emerge de sua estrutura tridimensional, que, por sua vez, é determinada pela sequência de aminoácidos e pelas condições físico-químicas do ambiente. Essa cadeia de determinantes — sequência → estrutura → dinâmica → função — define o modo como a informação genética é convertida em atividade biológica.
O primeiro princípio fundamental é o da complementaridade estrutural. Proteínas interagem com ligantes — substratos, cofatores, íons ou outras macromoléculas — por meio de superfícies que apresentam geometria e distribuição de cargas compatíveis com o parceiro de interação. Essa complementaridade não é absoluta nem rígida; ela é suficientemente específica para garantir seletividade, mas flexível o bastante para permitir ajustes conformacionais. O conceito clássico de “chave-fechadura” evoluiu para modelos mais realistas, nos quais a estrutura proteica pode adaptar-se ao ligante, refinando a interação e aumentando sua eficiência [Figura].
Um segundo princípio é o da organização espacial de grupos funcionais. A atividade de uma proteína depende da posição relativa de determinados resíduos de aminoácidos no espaço tridimensional. Em enzimas, por exemplo, o sítio ativo reúne grupos químicos capazes de estabilizar estados de transição, transferir prótons ou elétrons e posicionar substratos com precisão. Esses resíduos frequentemente não estão próximos na sequência primária, mas convergem espacialmente durante o dobramento. Assim, a função não está codificada apenas na presença de determinados aminoácidos, mas na sua disposição tridimensional.
A dinâmica conformacional constitui o terceiro princípio essencial. Proteínas não operam como estruturas estáticas; sua função envolve transições entre diferentes estados conformacionais. Essas mudanças podem ser sutis, como rotações de cadeias laterais, ou mais amplas, envolvendo movimentos de domínios inteiros. A capacidade de alternar entre estados estruturais permite que proteínas realizem ciclos funcionais, como ligação e liberação de substratos, ativação e inativação enzimática, ou transmissão de sinais moleculares. Sem essa flexibilidade, a função biológica seria severamente limitada.
Outro princípio central é o da cooperação e comunicação intramolecular. Em proteínas com múltiplos sítios funcionais ou múltiplas subunidades, eventos em uma região podem influenciar outras regiões distantes. Esse fenômeno, frequentemente associado ao alosterismo, depende da propagação de mudanças conformacionais através da estrutura proteica. A arquitetura tridimensional atua, portanto, como uma rede de comunicação interna, permitindo integração de sinais e regulação fina da atividade.
A relação estrutura–função também é governada pelo princípio da otimização energética. A conformação nativa de uma proteína corresponde, em geral, a um estado de baixa energia livre, mas não necessariamente ao mínimo absoluto. Essa condição permite um equilíbrio entre estabilidade e flexibilidade, essencial para a função. Proteínas excessivamente estáveis podem tornar-se rígidas e perder atividade, enquanto proteínas pouco estáveis podem desnaturar ou agregar. A evolução seleciona sequências que atingem esse ponto ótimo, maximizando a eficiência funcional em condições fisiológicas.
A sensibilidade ao ambiente representa outro aspecto crítico dessa relação. Alterações em pH, temperatura, força iônica ou concentração de ligantes podem modificar a estrutura proteica e, consequentemente, sua função. Essa dependência permite que proteínas atuem como sensores moleculares, ajustando sua atividade em resposta a mudanças no meio. No contexto celular, essa propriedade é explorada em processos regulatórios, nos quais pequenas variações ambientais desencadeiam respostas bioquímicas amplificadas.
Do ponto de vista evolutivo, a relação estrutura–função é moldada por pressões seletivas que favorecem eficiência e adaptação. Regiões estruturais críticas tendem a ser altamente conservadas, enquanto regiões periféricas podem tolerar variações, permitindo a diversificação funcional. A reutilização de domínios estruturais em diferentes contextos evidencia que a natureza explora soluções já estabelecidas, recombinando módulos para gerar novas funções. Esse princípio explica a recorrência de motivos estruturais em proteínas com funções distintas.
A ruptura dessa relação, seja por mutações, modificações químicas ou condições ambientais adversas, pode levar à perda de função ou ao ganho de funções indesejadas. Muitas doenças têm origem em alterações estruturais que comprometem a atividade proteica ou promovem agregação anormal. Esses fenômenos reforçam que a integridade estrutural é indispensável para a manutenção da função biológica.
Portanto, a relação estrutura–função em proteínas não é um conceito isolado, mas um conjunto de princípios interdependentes que explicam como moléculas biológicas executam tarefas com precisão e eficiência. Compreender essa relação permite não apenas interpretar processos naturais, mas também projetar intervenções, como o desenvolvimento de fármacos, a engenharia de enzimas e a otimização de sistemas biotecnológicos. Em última análise, é nesse vínculo que se revela a lógica molecular da vida, onde forma, energia e movimento convergem para gerar função [Quadro].
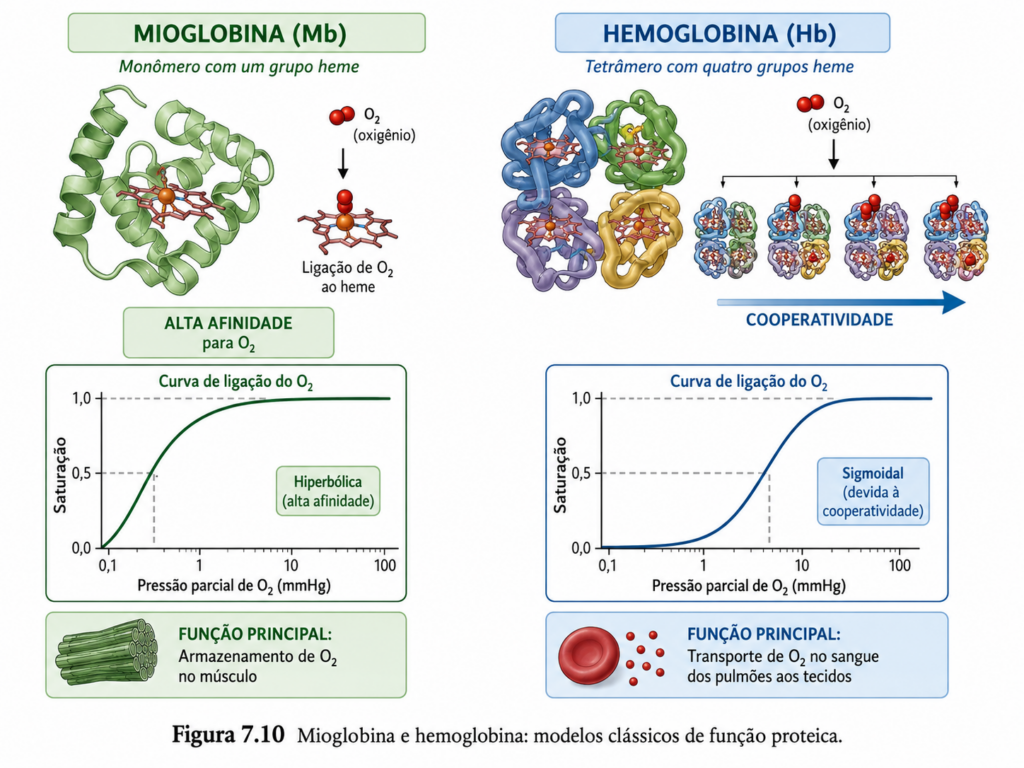
7.10 Mioglobina e hemoglobina: modelos clássicos de função proteica #
A compreensão da relação entre estrutura e função em proteínas alcança um de seus exemplos mais didáticos e completos no estudo da mioglobina e da hemoglobina. Essas duas proteínas, ambas especializadas na ligação ao oxigênio, ilustram como variações na organização estrutural — particularmente na presença ou ausência de estrutura quaternária — resultam em comportamentos funcionais profundamente distintos. Ao analisá-las, torna-se evidente que pequenas diferenças estruturais podem gerar respostas fisiológicas altamente adaptativas.
A mioglobina é uma proteína monomérica encontrada principalmente no tecido muscular, onde atua como reservatório de oxigênio. Sua estrutura é tipicamente globular, composta predominantemente por α-hélices organizadas de forma compacta ao redor de um grupo prostético central: o heme. Esse grupo contém um átomo de ferro (Fe²⁺) capaz de ligar-se reversivelmente ao oxigênio molecular. A interação entre o ferro e o oxigênio não ocorre de forma isolada; ela é modulada pelo ambiente proteico, que controla a afinidade e evita a oxidação do ferro para Fe³⁺, estado no qual a ligação ao oxigênio não seria possível [Figura].
A afinidade da mioglobina pelo oxigênio é relativamente alta, o que se reflete em uma curva de saturação hiperbólica. Isso significa que a proteína liga-se ao oxigênio de forma eficiente mesmo em baixas pressões parciais, característica compatível com sua função de armazenamento e liberação controlada em condições de baixa disponibilidade de oxigênio, como durante atividade muscular intensa. Nesse contexto, a mioglobina atua como um sistema de amortecimento, garantindo fornecimento contínuo de oxigênio às células musculares.
Em contraste, a hemoglobina é uma proteína tetramérica presente nas hemácias, responsável pelo transporte de oxigênio dos pulmões para os tecidos. Sua estrutura quaternária é composta por quatro subunidades (duas α e duas β em sua forma adulta), cada uma contendo um grupo heme semelhante ao da mioglobina. No entanto, a presença de múltiplas subunidades introduz um nível adicional de complexidade funcional: a cooperatividade na ligação ao oxigênio.
A hemoglobina exibe uma curva de saturação sigmoide, refletindo o fenômeno de cooperatividade positiva. A ligação de uma molécula de oxigênio a uma subunidade induz uma mudança conformacional que aumenta a afinidade das demais subunidades pelo oxigênio. Esse comportamento permite que a hemoglobina seja altamente eficiente tanto na captação de oxigênio em ambientes com alta pressão parcial (como os pulmões) quanto na liberação em tecidos com menor disponibilidade de oxigênio. Essa transição entre estados de baixa afinidade (estado T, tense) e alta afinidade (estado R, relaxed) é mediada por rearranjos estruturais coordenados entre as subunidades [Figura].
Além da cooperatividade, a hemoglobina é um exemplo clássico de regulação alostérica. Sua afinidade pelo oxigênio é modulada por diversos fatores fisiológicos, como pH, concentração de dióxido de carbono e presença de moduladores como o 2,3-bisfosfoglicerato (2,3-BPG). O chamado efeito Bohr descreve a diminuição da afinidade da hemoglobina pelo oxigênio em condições de pH mais baixo e maior concentração de CO₂, facilitando a liberação de oxigênio em tecidos metabolicamente ativos. O 2,3-BPG, por sua vez, liga-se à hemoglobina estabilizando o estado T, reduzindo a afinidade pelo oxigênio e favorecendo sua liberação [Quadro].
Do ponto de vista estrutural, tanto a mioglobina quanto a hemoglobina compartilham o mesmo motivo básico de dobramento — o “globin fold” — evidenciando uma origem evolutiva comum. No entanto, a organização quaternária da hemoglobina introduz propriedades emergentes que não estão presentes na mioglobina. Esse contraste demonstra como a evolução pode reutilizar um mesmo módulo estrutural, adaptando-o a diferentes funções por meio de alterações na organização molecular.
A relevância fisiológica dessas proteínas torna-se ainda mais evidente quando se consideram alterações estruturais decorrentes de mutações. A hemoglobina S, associada à anemia falciforme, resulta da substituição de um único aminoácido na cadeia β, levando à formação de agregados proteicos sob condições de baixa oxigenação. Esse fenômeno compromete a forma e a funcionalidade das hemácias, ilustrando de maneira direta como pequenas alterações na estrutura primária podem impactar drasticamente a função biológica.
Portanto, mioglobina e hemoglobina constituem modelos paradigmáticos para o estudo da bioquímica das proteínas. Elas demonstram, de forma integrada, como níveis estruturais distintos — desde o dobramento individual até a organização quaternária — influenciam propriedades funcionais como afinidade, cooperatividade e regulação. Mais do que exemplos específicos, essas proteínas sintetizam os princípios fundamentais que governam o comportamento das macromoléculas biológicas, conectando estrutura, dinâmica e função em um sistema altamente eficiente e adaptativo [Figura].
7.11 Cooperatividade e alosterismo em proteínas #
A capacidade de proteínas modularem sua atividade em resposta a mudanças no ambiente ou à ligação de ligantes é um dos pilares da regulação bioquímica. Dois conceitos centrais explicam esse comportamento: cooperatividade e alosterismo. Ambos derivam diretamente da organização estrutural das proteínas, especialmente daquelas que apresentam múltiplos sítios de ligação ou múltiplas subunidades, e refletem a habilidade dessas macromoléculas de integrar sinais moleculares de forma dinâmica e eficiente.
A cooperatividade refere-se ao fenômeno pelo qual a ligação de um ligante a uma proteína influencia a afinidade de outros sítios de ligação na mesma molécula. Quando essa influência aumenta a afinidade pelos ligantes subsequentes, temos a chamada cooperatividade positiva; quando a reduz, trata-se de cooperatividade negativa. Esse comportamento é característico de proteínas oligoméricas e resulta de mudanças conformacionais que se propagam entre as subunidades, alterando o estado estrutural do conjunto.
Do ponto de vista experimental, a cooperatividade é evidenciada por curvas de saturação sigmoides, em contraste com a forma hiperbólica observada em sistemas não cooperativos. Essa resposta sigmoide permite uma transição mais abrupta entre estados de baixa e alta saturação, tornando a proteína particularmente sensível a variações na concentração do ligante. Esse tipo de comportamento é altamente vantajoso em sistemas fisiológicos que exigem respostas rápidas e ajustáveis, como o transporte de oxigênio pela hemoglobina [Figura].
A base molecular da cooperatividade reside na existência de múltiplos estados conformacionais interconvertíveis. Dois modelos clássicos descrevem esse comportamento. No modelo concertado (ou modelo MWC), todas as subunidades de uma proteína alternam simultaneamente entre estados de baixa afinidade (estado T) e alta afinidade (estado R). A ligação do ligante estabiliza o estado R, deslocando o equilíbrio em sua direção. Já no modelo sequencial (ou modelo KNF), a ligação do ligante a uma subunidade induz uma mudança conformacional local que, progressivamente, influencia as subunidades vizinhas. Embora distintos em sua abordagem, ambos os modelos capturam aspectos reais do comportamento cooperativo, e proteínas biológicas frequentemente exibem características intermediárias entre eles [Quadro].
O alosterismo, por sua vez, refere-se à regulação da atividade proteica por meio da ligação de moléculas em sítios distintos do sítio ativo, denominados sítios alostéricos. Essa ligação provoca mudanças conformacionais que alteram a atividade da proteína, podendo ativá-la ou inibi-la. Diferentemente da cooperatividade, que envolve múltiplos sítios para o mesmo ligante, o alosterismo geralmente envolve ligantes diferentes que modulam a função da proteína.
A base estrutural do alosterismo é a capacidade da proteína de transmitir mudanças conformacionais ao longo de sua estrutura. A ligação de um efetor alostérico em uma região específica desencadeia rearranjos estruturais que afetam regiões distantes, incluindo o sítio ativo. Essa comunicação intramolecular depende da conectividade estrutural da proteína, que atua como uma rede de transmissão de sinais. Assim, proteínas alostéricas funcionam como dispositivos moleculares capazes de integrar múltiplos estímulos e ajustar sua atividade de forma coordenada.
Um exemplo clássico de alosterismo é a regulação enzimática em vias metabólicas. Muitas enzimas-chave são moduladas por produtos finais da via, em um mecanismo de retroalimentação negativa. Nesse contexto, o produto final atua como um inibidor alostérico, reduzindo a atividade da enzima inicial e evitando a produção excessiva. Esse tipo de controle é essencial para a homeostase celular, garantindo eficiência e economia de recursos metabólicos.
A interação entre cooperatividade e alosterismo amplia significativamente a capacidade regulatória das proteínas. Em muitos casos, proteínas exibem ambos os comportamentos simultaneamente, permitindo respostas altamente sensíveis e moduláveis. A presença de múltiplos ligantes, cada um influenciando a estrutura de maneira distinta, cria um sistema de regulação multifatorial, no qual a atividade proteica resulta da integração de diversos sinais bioquímicos.
Do ponto de vista termodinâmico, esses fenômenos refletem mudanças no equilíbrio entre diferentes estados conformacionais da proteína. Ligantes e efetores não criam novos estados, mas estabilizam estados já existentes, deslocando o equilíbrio em favor de conformações específicas. Essa perspectiva reforça a ideia de que proteínas existem como ensembles dinâmicos, e que sua função depende da distribuição relativa desses estados.
Alterações na cooperatividade ou no alosterismo podem ter consequências fisiológicas significativas. Mutações que afetam a comunicação entre subunidades ou a estabilidade de estados conformacionais podem comprometer a regulação proteica, levando a disfunções metabólicas ou doenças. Por outro lado, esses mecanismos também representam alvos estratégicos para intervenção farmacológica, uma vez que moduladores alostéricos podem ajustar a atividade de proteínas de forma mais específica do que inibidores competitivos.
Portanto, cooperatividade e alosterismo constituem mecanismos fundamentais pelos quais proteínas transcendem o comportamento passivo e assumem papel ativo na regulação dos sistemas biológicos. Eles exemplificam como a estrutura tridimensional e a dinâmica conformacional permitem que proteínas operem como sensores, integradores e moduladores de processos bioquímicos complexos, conectando diretamente a arquitetura molecular à regulação funcional [Figura].
7.12 Modulação funcional por ligantes e fatores ambientais (pH, temperatura, íons) #
A atividade de uma proteína não é determinada exclusivamente por sua estrutura intrínseca, mas também pelas condições do meio em que está inserida. Em sistemas biológicos, proteínas operam em ambientes dinâmicos, nos quais variáveis físico-químicas como pH, temperatura, força iônica e concentração de ligantes variam continuamente. A modulação funcional por esses fatores não representa uma perturbação externa, mas um componente essencial da regulação biológica, permitindo que proteínas ajustem sua atividade de forma precisa às demandas fisiológicas.
A interação com ligantes constitui o mecanismo mais direto de modulação funcional. Ligantes podem atuar como substratos, cofatores, inibidores ou moduladores alostéricos, influenciando a atividade proteica por meio de mudanças conformacionais. A ligação de um ligante pode estabilizar estados específicos da proteína, alterando sua afinidade por outros ligantes ou sua capacidade catalítica. Esse processo está profundamente relacionado à dinâmica conformacional, na qual a proteína alterna entre diferentes estados estruturais e o ligante atua como um fator de seleção ou indução dessas conformações [Figura].
O pH é um dos fatores ambientais mais críticos na regulação da atividade proteica. Ele influencia diretamente o estado de ionização dos grupos funcionais presentes nas cadeias laterais dos aminoácidos, especialmente aqueles com caráter ácido ou básico. Alterações no pH podem modificar cargas elétricas, afetando interações eletrostáticas, ligações de hidrogênio e a própria estrutura tridimensional da proteína. Em enzimas, o pH pode alterar a protonação de resíduos no sítio ativo, comprometendo mecanismos catalíticos que dependem de transferências de prótons. Por essa razão, muitas proteínas apresentam um intervalo ótimo de pH no qual sua atividade é maximizada, refletindo uma adaptação às condições fisiológicas específicas de seu ambiente [Quadro].
A temperatura exerce influência tanto sobre a cinética quanto sobre a estabilidade estrutural das proteínas. O aumento da temperatura tende a acelerar reações químicas, elevando a taxa de colisões moleculares e, consequentemente, a velocidade catalítica. No entanto, temperaturas elevadas também aumentam a energia cinética das moléculas, podendo desestabilizar interações não covalentes que mantêm a estrutura proteica. Esse efeito pode levar à desnaturação, caracterizada pela perda da conformação tridimensional e da função biológica. Por outro lado, temperaturas muito baixas reduzem a flexibilidade molecular, podendo limitar a dinâmica conformacional necessária para a atividade. Assim, cada proteína apresenta uma faixa de temperatura ótima que equilibra eficiência catalítica e estabilidade estrutural [Figura].
Os íons e a força iônica do meio também desempenham papel relevante na modulação funcional. Íons podem interagir diretamente com proteínas, estabilizando estruturas ou participando de mecanismos catalíticos. Metais como zinco, magnésio e ferro frequentemente atuam como cofatores enzimáticos, facilitando reações químicas por meio de mecanismos de polarização de ligações ou estabilização de intermediários. Além disso, a presença de íons em solução influencia as interações eletrostáticas entre resíduos carregados, podendo fortalecer ou enfraquecer pontes salinas e afetar a estabilidade global da proteína.
A concentração de ligantes moduladores, como o 2,3-bisfosfoglicerato na hemoglobina, exemplifica como fatores químicos específicos podem ajustar a função proteica de maneira refinada. Esses moduladores não participam diretamente da reação principal, mas alteram a afinidade da proteína por seus ligantes principais, permitindo adaptação a diferentes condições fisiológicas. Esse tipo de regulação é particularmente importante em sistemas que exigem respostas rápidas a mudanças ambientais, como o transporte de gases ou o controle metabólico.
Outro aspecto relevante é a interação entre esses fatores. Em sistemas biológicos, pH, temperatura e concentração de íons não atuam isoladamente, mas de forma integrada. Alterações em um parâmetro podem influenciar a sensibilidade da proteína a outros, criando um sistema de regulação multifatorial. Por exemplo, mudanças no pH podem alterar a afinidade por íons metálicos, enquanto variações de temperatura podem modificar a estabilidade de complexos proteína-ligante.
Do ponto de vista estrutural, a modulação por fatores ambientais reflete a natureza delicadamente equilibrada das interações que mantêm a conformação proteica. Pequenas perturbações podem deslocar o equilíbrio entre estados conformacionais, alterando a função sem necessariamente causar desnaturação completa. Essa sensibilidade é explorada biologicamente para regular processos celulares, mas também representa um ponto de vulnerabilidade, uma vez que condições extremas podem comprometer a integridade proteica.
Em aplicações práticas, especialmente em contextos industriais e agrícolas, o controle desses fatores é fundamental para otimizar a atividade de proteínas, como enzimas utilizadas em processos biotecnológicos. Ajustes finos de pH, temperatura e composição iônica permitem maximizar rendimento e eficiência, demonstrando a importância de compreender esses princípios além do contexto puramente teórico.
Portanto, a modulação funcional por ligantes e fatores ambientais evidencia que proteínas não são entidades isoladas, mas sistemas sensíveis ao contexto em que operam. Sua atividade resulta da interação contínua entre estrutura interna e condições externas, permitindo respostas adaptativas que são essenciais para a manutenção da vida. Essa capacidade de ajuste dinâmico reforça a ideia de que a função proteica é um fenômeno emergente, dependente tanto da arquitetura molecular quanto do ambiente em que se manifesta [Figura].
7.13 Desnaturação, renaturação e estabilidade proteica #
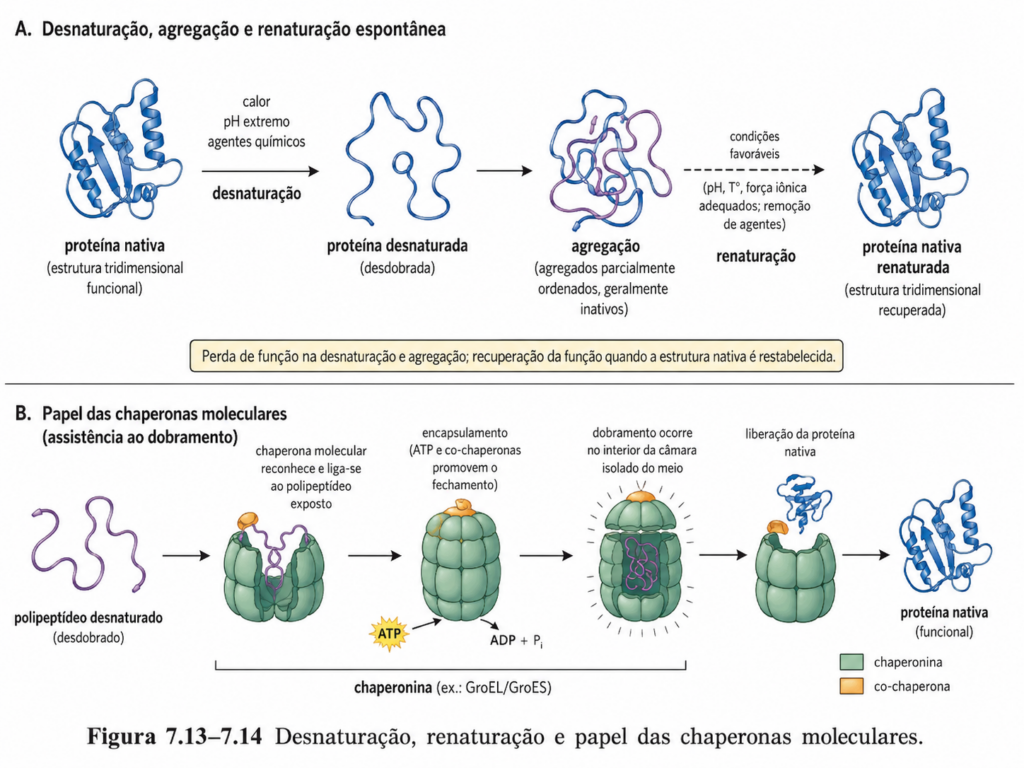
A estrutura tridimensional das proteínas, embora altamente organizada e funcionalmente precisa, não é intrinsecamente permanente. Ela depende de um conjunto de interações químicas delicadamente equilibradas, suscetíveis a perturbações externas. A desnaturação corresponde ao processo pelo qual essa organização estrutural é perdida, levando à desestabilização da conformação nativa e, consequentemente, à perda da função biológica. Esse fenômeno evidencia que a atividade proteica está diretamente vinculada à integridade estrutural, e não apenas à composição química da molécula.
Durante a desnaturação, as estruturas secundária, terciária e quaternária são desorganizadas, enquanto a estrutura primária — as ligações peptídicas — permanece, em geral, intacta. Isso ocorre porque as interações responsáveis pela manutenção da estrutura tridimensional são predominantemente não covalentes, como ligações de hidrogênio, interações hidrofóbicas e forças eletrostáticas, que são relativamente sensíveis a alterações ambientais. A ruptura dessas interações resulta na exposição de regiões internas da proteína, frequentemente hidrofóbicas, ao meio aquoso, alterando drasticamente suas propriedades físico-químicas [Figura].
Diversos agentes podem induzir desnaturação. O aumento da temperatura é um dos mais comuns, pois eleva a energia cinética das moléculas, superando as forças estabilizadoras e promovendo a desorganização estrutural. Alterações extremas de pH modificam o estado de ionização dos grupos funcionais, rompendo interações eletrostáticas e ligações de hidrogênio. Agentes químicos, como ureia e cloreto de guanidínio, interferem diretamente nas interações não covalentes, enquanto detergentes podem solubilizar regiões hidrofóbicas, desestabilizando o núcleo proteico. Além disso, solventes orgânicos e metais pesados também podem comprometer a estrutura, seja por interação direta com resíduos específicos, seja por alteração do ambiente químico.
Um aspecto relevante da desnaturação é que, em muitos casos, ela é acompanhada por agregação proteica. Ao perder sua estrutura nativa, regiões hidrofóbicas anteriormente protegidas tornam-se expostas, favorecendo interações entre diferentes moléculas de proteína. Esse processo leva à formação de agregados insolúveis, frequentemente irreversíveis, que impedem a recuperação da estrutura original. Esse fenômeno tem implicações biológicas importantes, estando associado a diversas doenças caracterizadas pelo acúmulo de proteínas mal dobradas.
Em contraste com a desnaturação, a renaturação refere-se à capacidade de uma proteína recuperar sua estrutura tridimensional nativa após a remoção do agente desnaturante. Esse processo é possível porque a informação necessária para o dobramento está contida na própria sequência de aminoácidos. Experimentos clássicos demonstraram que, sob condições adequadas, proteínas desnaturadas podem retornar espontaneamente ao seu estado funcional, reforçando o princípio de que a estrutura primária determina a estrutura tridimensional [Quadro].
No entanto, a renaturação não é universalmente eficiente. Em ambientes celulares, onde a concentração de macromoléculas é elevada, a probabilidade de interações incorretas e agregação é significativa. Por essa razão, células utilizam chaperonas moleculares, proteínas especializadas que auxiliam no dobramento correto, prevenindo agregações indesejadas. Essas chaperonas não determinam a estrutura final, mas fornecem um ambiente controlado para que o dobramento ocorra de forma eficiente.
A estabilidade proteica deve ser entendida como a resistência da proteína à desnaturação, refletindo o equilíbrio entre o estado nativo e estados desenovelados. Esse equilíbrio é descrito em termos termodinâmicos, considerando a diferença de energia livre entre esses estados. Proteínas estáveis apresentam uma diferença de energia livre favorável ao estado nativo, mas essa diferença não é excessivamente grande, o que permite certa flexibilidade estrutural necessária à função. Assim, estabilidade não implica rigidez absoluta, mas sim um balanço entre integridade estrutural e dinâmica conformacional.
Fatores intrínsecos e extrínsecos influenciam essa estabilidade. A composição de aminoácidos, a presença de pontes dissulfeto, o grau de empacotamento interno e a organização de domínios contribuem para a resistência estrutural. Por outro lado, condições ambientais, como temperatura, pH e presença de solutos, modulam continuamente esse equilíbrio. Em organismos adaptados a ambientes extremos, como termófilos, proteínas apresentam adaptações estruturais específicas que aumentam sua estabilidade térmica, evidenciando a capacidade evolutiva de otimizar essas propriedades.
Do ponto de vista funcional, a desnaturação representa a perda de atividade, enquanto a renaturação, quando possível, restaura a função. Esse ciclo ressalta que a atividade biológica não é uma propriedade permanente da molécula, mas depende de sua conformação estrutural. Em aplicações biotecnológicas, controlar esses processos é essencial para preservar a atividade de enzimas e outras proteínas, seja durante armazenamento, processamento ou uso industrial.
Portanto, desnaturação, renaturação e estabilidade proteica constituem aspectos fundamentais da bioquímica das proteínas, evidenciando a natureza dinâmica e sensível dessas moléculas. Eles demonstram que a função biológica é inseparável da estrutura tridimensional e que essa estrutura, embora robusta, está em constante interação com o ambiente. Compreender esses processos é essencial para interpretar tanto fenômenos fisiológicos quanto aplicações tecnológicas e patológicas relacionadas às proteínas [Figura].
7.14 Dobramento proteico e papel das chaperonas moleculares #
O dobramento proteico é o processo pelo qual uma cadeia polipeptídica recém-sintetizada adquire sua conformação tridimensional funcional. Embora a informação necessária para esse processo esteja contida na sequência de aminoácidos, a transição entre uma cadeia linear e uma estrutura altamente organizada envolve uma série de eventos coordenados, guiados por princípios termodinâmicos e modulados pelo ambiente celular. Esse processo não é apenas uma etapa intermediária, mas um determinante crítico da função biológica das proteínas.
Do ponto de vista energético, o dobramento pode ser descrito como a progressão de uma proteína ao longo de uma paisagem de energia livre, frequentemente representada como um funil. Nesse modelo, múltiplos estados conformacionais de alta energia convergem progressivamente para um conjunto de estados de menor energia, correspondentes à conformação nativa. Essa trajetória não é única nem linear; diferentes caminhos podem levar ao mesmo estado final, passando por intermediários parcialmente dobrados. A eficiência desse processo resolve o chamado paradoxo de Levinthal, que aponta a improbabilidade de uma busca aleatória por todas as conformações possíveis [Figura].
Durante o dobramento, a formação de estruturas secundárias ocorre rapidamente, seguida pelo estabelecimento de interações de longo alcance que definem a estrutura terciária. O colapso hidrofóbico desempenha papel central, promovendo a formação de um núcleo interno compacto. No entanto, esse processo não está isento de riscos. Intermediários mal dobrados podem expor regiões hidrofóbicas, favorecendo interações incorretas e levando à agregação proteica. Em condições ideais, a proteína atinge sua conformação nativa de forma espontânea; entretanto, no ambiente celular, onde a concentração de macromoléculas é elevada, a probabilidade de erros aumenta significativamente.
É nesse contexto que atuam as chaperonas moleculares, proteínas especializadas que auxiliam no dobramento correto de outras proteínas. Essas moléculas não fazem parte da estrutura final da proteína-alvo, nem determinam diretamente sua conformação, mas fornecem um ambiente controlado que reduz a ocorrência de interações incorretas. Sua função principal é prevenir a agregação e facilitar a progressão da proteína ao longo da paisagem energética em direção ao estado nativo.
Entre as chaperonas mais estudadas estão as proteínas da família Hsp70, que se ligam a segmentos hidrofóbicos expostos em polipeptídeos nascentes, impedindo interações indevidas. Esse processo é regulado por ciclos de ligação e liberação dependentes de ATP, permitindo que a proteína-alvo explore diferentes conformações até atingir um estado estável. Outro sistema importante é o das chaperoninas, como o complexo GroEL/GroES em bactérias, que formam uma cavidade isolada onde o dobramento pode ocorrer de forma protegida. Nesse ambiente, a proteína é temporariamente encapsulada, reduzindo a interferência do meio externo e aumentando a probabilidade de dobramento correto [Figura].
O papel das chaperonas torna-se ainda mais crítico em situações de estresse celular, como aumento de temperatura, presença de toxinas ou condições oxidativas. Nessas circunstâncias, proteínas previamente dobradas podem sofrer desnaturação parcial, exigindo mecanismos de reparo ou redobramento. A expressão de chaperonas é frequentemente induzida nesses contextos, caracterizando a chamada resposta ao choque térmico. Esse sistema de controle evidencia que o dobramento proteico não é um evento isolado, mas um processo continuamente monitorado pela célula.
A falha nos mecanismos de dobramento e controle de qualidade pode levar ao acúmulo de proteínas mal dobradas ou agregadas, fenômeno associado a diversas doenças. Em patologias neurodegenerativas, como Alzheimer e Parkinson, agregados proteicos insolúveis interferem na função celular, destacando a importância do equilíbrio entre dobramento correto, degradação e manutenção da proteostase. Nesse contexto, sistemas celulares como o proteassomo e a autofagia atuam na remoção de proteínas defeituosas, complementando a ação das chaperonas.
Do ponto de vista evolutivo, a existência de chaperonas permitiu a expansão da complexidade proteica. Proteínas maiores e mais complexas, com múltiplos domínios, são mais propensas a erros de dobramento. A presença de sistemas auxiliares de dobramento reduziu essa limitação, possibilitando o surgimento de novas funções e aumentando a diversidade estrutural das proteínas. Assim, chaperonas não apenas mantêm a integridade funcional, mas também contribuem para a inovação evolutiva.
Em aplicações biotecnológicas, o controle do dobramento proteico é um desafio central, especialmente na produção de proteínas recombinantes. A expressão heteróloga frequentemente resulta em proteínas mal dobradas ou agregadas, exigindo estratégias que envolvem coexpressão de chaperonas, otimização de condições de cultivo ou renaturação in vitro. A compreensão dos mecanismos de dobramento e da ação das chaperonas é, portanto, essencial para o desenvolvimento de processos eficientes e para a produção de proteínas funcionais em escala industrial.
Portanto, o dobramento proteico e a atuação das chaperonas moleculares constituem um sistema integrado que garante a funcionalidade das proteínas em ambientes celulares complexos. Esse processo evidencia que a estrutura proteica não é apenas resultado de propriedades intrínsecas da sequência, mas também de um contexto biológico que orienta, protege e corrige o caminho até a conformação nativa. A função proteica, nesse sentido, emerge da interação entre informação molecular e mecanismos celulares de controle, consolidando a proteína como uma unidade dinâmica e regulada da vida [Quadro].
7.15 Proteínas em sistemas aplicados: agricultura, indústria e biotecnologia #
A compreensão das proteínas ultrapassa o domínio descritivo da bioquímica estrutural e alcança aplicações diretas em sistemas produtivos, tecnológicos e ambientais. Em contextos aplicados, proteínas deixam de ser apenas objetos de estudo e passam a atuar como ferramentas estratégicas, capazes de aumentar eficiência, reduzir custos e promover inovação em múltiplos setores. A relação entre estrutura, estabilidade e função, discutida nos tópicos anteriores, torna-se aqui um instrumento de intervenção e otimização.
No contexto da agricultura, proteínas desempenham papel central tanto no metabolismo vegetal quanto na interface com o ambiente. Enzimas regulam processos como fotossíntese, assimilação de nitrogênio, síntese de biomassa e resposta a estresses abióticos e bióticos. A manipulação dessas proteínas, seja por melhoramento genético ou engenharia molecular, permite aumentar a produtividade e a resiliência das culturas. Por exemplo, a modulação da atividade de enzimas envolvidas na fixação de carbono pode impactar diretamente a eficiência fotossintética, enquanto proteínas relacionadas à resposta ao estresse hídrico contribuem para a adaptação ao semiárido [Figura].
Além disso, proteínas são fundamentais na nutrição vegetal e animal. A qualidade proteica de culturas agrícolas influencia diretamente o valor nutricional dos alimentos, sendo alvo de programas de biofortificação. No manejo agrícola, enzimas também são utilizadas como indicadores bioquímicos da saúde do solo, refletindo atividade microbiana e ciclos de nutrientes. Esse uso conecta bioquímica molecular a práticas de campo, permitindo monitoramento e tomada de decisão baseada em parâmetros funcionais.
Na indústria, proteínas — especialmente enzimas — são amplamente exploradas como catalisadores biológicos. Sua alta especificidade e eficiência permitem a realização de reações químicas em condições mais brandas, reduzindo consumo energético e geração de resíduos. Setores como alimentos, detergentes, farmacêutico e têxtil utilizam enzimas para processos como hidrólise, síntese de compostos e modificação de biomoléculas. A estabilidade proteica, discutida anteriormente, torna-se um fator crítico, uma vez que condições industriais frequentemente envolvem variações de temperatura, pH e presença de solventes.
A engenharia de proteínas tem permitido o desenvolvimento de enzimas com propriedades otimizadas para aplicações específicas. Por meio de mutagênese dirigida ou evolução dirigida, é possível alterar a estrutura primária e, consequentemente, ajustar estabilidade, atividade catalítica e especificidade. Esse processo exemplifica, de forma prática, o princípio de que pequenas alterações estruturais podem gerar mudanças funcionais significativas. A capacidade de projetar proteínas com características desejadas representa um dos avanços mais relevantes da biotecnologia moderna.
No campo da biotecnologia, proteínas assumem papel ainda mais abrangente. Elas são utilizadas como biofármacos, incluindo anticorpos monoclonais, hormônios e enzimas terapêuticas. A eficácia desses produtos depende diretamente de sua estrutura correta e estabilidade, exigindo controle rigoroso durante produção, purificação e armazenamento. Proteínas também são utilizadas como ferramentas analíticas, como em biossensores capazes de detectar moléculas específicas com alta sensibilidade.
Outro avanço relevante é o uso de proteínas em sistemas de edição genética e biologia sintética. Enzimas como nucleases programáveis permitem modificar o genoma de organismos com precisão, abrindo possibilidades para desenvolvimento de culturas mais produtivas, resistentes ou adaptadas a condições específicas. Nesse contexto, a compreensão da estrutura e da dinâmica proteica é essencial para garantir especificidade e minimizar efeitos indesejados.
A aplicação de proteínas também se estende à sustentabilidade e ao desenvolvimento de tecnologias verdes. Enzimas são utilizadas em processos de biorremediação, degradando poluentes e contribuindo para recuperação ambiental. Na produção de biocombustíveis, proteínas participam da conversão de biomassa em energia, integrando bioquímica a soluções energéticas renováveis. Esses exemplos demonstram como princípios moleculares podem ser traduzidos em impacto ambiental e econômico.
Um ponto crítico em sistemas aplicados é o controle da estabilidade funcional das proteínas. Condições não fisiológicas podem levar à desnaturação ou perda de atividade, exigindo estratégias como imobilização enzimática, modificação química ou uso de aditivos estabilizantes. Essas abordagens visam prolongar a vida útil das proteínas e aumentar sua eficiência em processos contínuos. A engenharia de estabilidade, portanto, torna-se tão relevante quanto a engenharia de atividade.
Do ponto de vista estratégico, a aplicação de proteínas representa uma convergência entre ciência básica e inovação tecnológica. A capacidade de manipular estrutura e função permite desenvolver soluções sob medida para problemas específicos, seja na produção de alimentos, no controle de doenças ou na geração de novos materiais. Essa integração exige não apenas conhecimento bioquímico, mas também visão sistêmica, capaz de conectar níveis moleculares a demandas reais.
Portanto, as proteínas em sistemas aplicados exemplificam a transição da bioquímica como ciência descritiva para uma bioquímica operacional e estratégica. Elas demonstram que compreender estrutura, dinâmica e função não é um fim em si mesmo, mas um meio para intervir, otimizar e inovar. Em um cenário de crescente demanda por eficiência e sustentabilidade, a exploração racional das proteínas posiciona-se como um dos principais vetores de transformação tecnológica e produtiva [Quadro].
Perguntas de estudo dirigido — Capítulo 7: Estrutura e atividade de proteínas #
- Explique como a sequência primária de aminoácidos influencia o dobramento tridimensional de uma proteína.
Na resposta, relacione propriedades das cadeias laterais, como polaridade, carga, hidrofobicidade, presença de glicina, prolina e cisteína, com a formação da estrutura final. - Compare estrutura secundária, terciária e quaternária das proteínas.
Indique quais interações químicas predominam em cada nível estrutural e explique por que esses níveis não devem ser interpretados como etapas totalmente independentes. - Diferencie proteínas globulares e proteínas fibrosas quanto à estrutura, solubilidade e função biológica.
Use exemplos como enzimas, hemoglobina, colágeno e queratina para justificar a comparação. - Explique por que mioglobina e hemoglobina apresentam comportamentos distintos na ligação ao oxigênio.
Relacione a estrutura monomérica da mioglobina, a estrutura tetramérica da hemoglobina, a cooperatividade e as curvas de saturação hiperbólica e sigmoide. - Descreva o processo de desnaturação proteica e discuta em que condições a renaturação pode ocorrer.
Inclua na resposta os efeitos de temperatura, pH, agentes químicos, exposição de regiões hidrofóbicas, agregação e participação das chaperonas moleculares.
Pergunta para estimular pesquisa #
- Como a engenharia de proteínas pode ser usada para desenvolver enzimas mais estáveis e eficientes para aplicações agrícolas, industriais ou biotecnológicas?
Pesquise um exemplo real de enzima modificada por engenharia de proteínas ou evolução dirigida e explique: qual proteína foi alterada, qual característica foi melhorada, qual técnica foi usada e qual aplicação prática resultou dessa modificação.
Referências #
NELSON, David L.; COX, Michael M. Princípios de bioquímica de Lehninger. 6. ed. Porto Alegre: Artmed, 2014.
VOET, Donald; VOET, Judith G.; PRATT, Charlotte W. Fundamentos de bioquímica: a vida em nível molecular. 4. ed. Porto Alegre: Artmed, 2014.
STRYER, Lubert; BERG, Jeremy M.; TYMOCZKO, John L.; GATTO JR., Gregory J. Bioquímica. 8. ed. Rio de Janeiro: Guanabara Koogan, 2014.
MURRAY, Robert K. et al. Bioquímica ilustrada de Harper. 30. ed. Porto Alegre: AMGH, 2017.



