As proteínas são os principais executores das funções celulares. Este capítulo introduz os aminoácidos como unidades básicas e explora a formação da ligação peptídica. A organização estrutural das proteínas é apresentada em diferentes níveis, evidenciando como pequenas variações estruturais impactam diretamente a função. Essa base é essencial para compreender a complexidade funcional das biomoléculas.
6.1 Estrutura geral dos aminoácidos #
A compreensão da bioquímica das proteínas começa inevitavelmente pela análise detalhada de suas unidades fundamentais: os aminoácidos. Essas moléculas, embora estruturalmente simples quando consideradas isoladamente, constituem um repertório químico extraordinariamente versátil, capaz de sustentar a diversidade funcional observada nos sistemas biológicos. Cada aminoácido é organizado em torno de um carbono central, denominado carbono α, ao qual estão ligados quatro substituintes distintos: um grupo amino, um grupo carboxila, um átomo de hidrogênio e uma cadeia lateral variável, convencionalmente designada como grupo R. Essa arquitetura confere aos aminoácidos uma dualidade essencial: uma base estrutural comum combinada com uma diversidade química determinada pelas propriedades específicas de suas cadeias laterais.
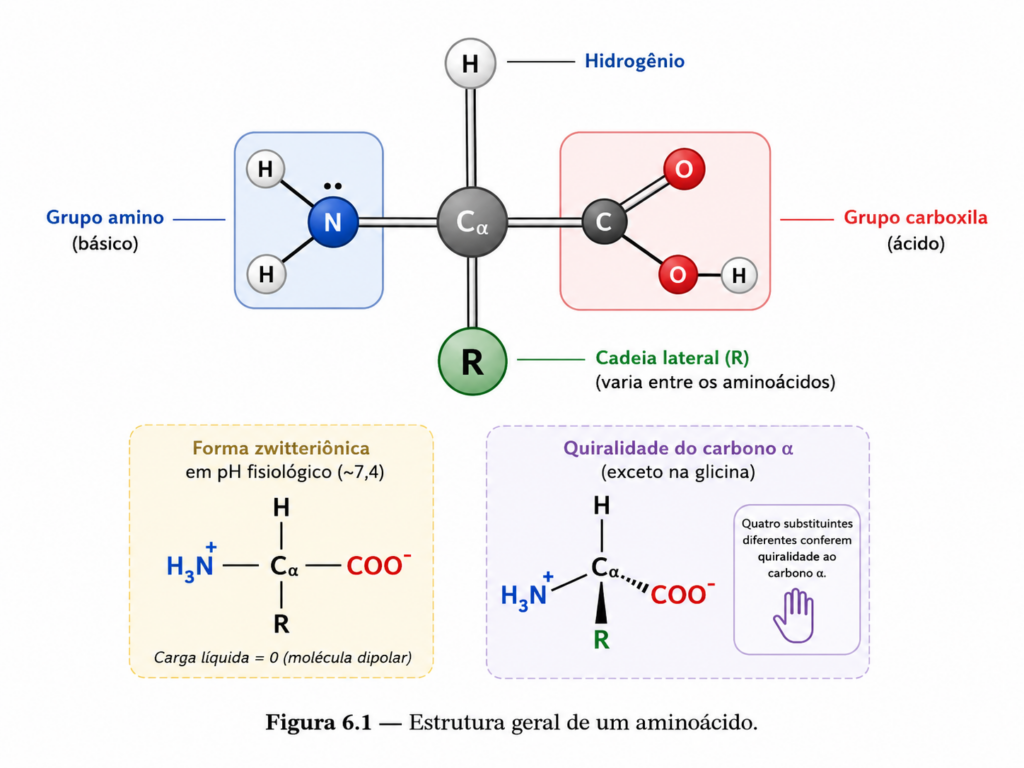
Em solução aquosa, sob condições fisiológicas, os aminoácidos não existem predominantemente em suas formas neutras, mas sim como íons dipolares, conhecidos como zwitterions. Nessa configuração, o grupo amino encontra-se protonado (–NH₃⁺), enquanto o grupo carboxila está desprotonado (–COO⁻), resultando em uma molécula globalmente neutra, porém eletricamente polar. Essa característica tem implicações diretas sobre a solubilidade, a reatividade e a interação dos aminoácidos com outras biomoléculas, sendo um dos fundamentos que explicam o comportamento das proteínas em ambientes biológicos complexos.
A natureza da cadeia lateral (R) é o principal determinante das propriedades químicas e físicas de cada aminoácido. Essa diversidade permite classificá-los em grupos funcionais, geralmente organizados em categorias como aminoácidos apolares, polares sem carga, carregados positivamente e carregados negativamente. Os aminoácidos apolares, como alanina, valina e leucina, apresentam cadeias laterais predominantemente hidrocarbonadas, o que favorece interações hidrofóbicas e sua tendência de se localizar no interior das proteínas globulares. Em contraste, aminoácidos polares e carregados, como serina, ácido aspártico e lisina, possuem grupos funcionais capazes de estabelecer interações com a água e com outras moléculas polares, desempenhando papéis cruciais em superfícies proteicas e sítios ativos.
Outro aspecto estrutural relevante é a quiralidade do carbono α. Com exceção da glicina, cuja cadeia lateral é um hidrogênio adicional, todos os aminoácidos apresentam quatro substituintes diferentes ligados ao carbono α, tornando-o um centro quiral. Como resultado, existem duas configurações possíveis, denominadas L e D. Nos sistemas biológicos, há uma predominância quase absoluta da configuração L, um fato que reflete a história evolutiva da vida e que tem profundas implicações na estrutura tridimensional das proteínas. A uniformidade estereoquímica permite a formação de estruturas regulares, como hélices e folhas β, que dependem de orientações espaciais consistentes ao longo da cadeia polipeptídica.
Além disso, os aminoácidos possuem propriedades ácido-base que são essenciais para sua função biológica. Cada molécula apresenta pelo menos dois grupos ionizáveis — o grupo amino e o grupo carboxila — e, em muitos casos, a própria cadeia lateral também pode ionizar. A capacidade de doar ou aceitar prótons depende do pH do meio e dos valores de pKa desses grupos. Essa característica confere aos aminoácidos um papel central na manutenção do equilíbrio ácido-base em sistemas biológicos, além de influenciar diretamente a estrutura e a atividade das proteínas. Pequenas variações de pH podem alterar o estado de ionização de resíduos específicos, modulando interações intramoleculares e, consequentemente, a conformação proteica.
A diversidade estrutural dos aminoácidos também se reflete em suas propriedades espectroscópicas e químicas específicas. Aminoácidos aromáticos, como fenilalanina, tirosina e triptofano, apresentam sistemas de elétrons π conjugados que absorvem luz na região ultravioleta, propriedade amplamente explorada na quantificação de proteínas [Tabela]. Outros, como a cisteína, contêm grupos sulfidrila capazes de formar ligações dissulfeto, fundamentais para a estabilização de estruturas tridimensionais em proteínas extracelulares. Esses detalhes estruturais, embora aparentemente pontuais, são determinantes para a funcionalidade das proteínas e para sua adaptação a diferentes contextos fisiológicos.
Do ponto de vista evolutivo e funcional, o conjunto padrão de vinte aminoácidos proteicos representa uma solução química altamente otimizada. Esse repertório fornece um equilíbrio entre diversidade estrutural e eficiência biossintética, permitindo a construção de macromoléculas com propriedades específicas a partir de um conjunto relativamente limitado de blocos construtores. A organização desses aminoácidos em sequências definidas — que será explorada nos tópicos seguintes — é o que transforma essa diversidade potencial em função biológica concreta.
Assim, ao analisar a estrutura geral dos aminoácidos, torna-se evidente que sua importância transcende a simples condição de monômeros das proteínas. Eles constituem, na realidade, unidades químicas multifuncionais, cuja organização, propriedades e interações definem os princípios fundamentais que governam a estrutura e a função das proteínas nos sistemas vivos.
6.2 Propriedades químicas dos aminoácidos #
A análise das propriedades químicas dos aminoácidos revela um aspecto central da bioquímica estrutural: essas moléculas não são entidades estáticas, mas sistemas dinâmicos cujo comportamento depende diretamente do ambiente químico em que se encontram. Entre os fatores mais determinantes está o pH do meio, que regula o estado de ionização dos grupos funcionais presentes nos aminoácidos e, consequentemente, influencia sua reatividade, solubilidade e participação em interações moleculares.
Cada aminoácido contém, no mínimo, dois grupos ionizáveis — o grupo carboxila e o grupo amino — que podem doar ou aceitar prótons dependendo das condições do meio. O grupo carboxila, com caráter ácido, tende a perder um próton, formando o íon carboxilato (–COO⁻), enquanto o grupo amino, com caráter básico, tende a aceitar um próton, formando o grupo amônio (–NH₃⁺). A extensão dessas ionizações é descrita quantitativamente pelos valores de pKa, que representam o pH no qual metade das moléculas daquele grupo funcional está ionizada. Esses valores funcionam como parâmetros fundamentais para prever o comportamento dos aminoácidos em diferentes contextos fisiológicos.
Quando se considera a variação do pH, observa-se que os aminoácidos passam por diferentes estados de carga líquida. Em pH muito ácido, ambos os grupos encontram-se protonados, resultando em uma carga líquida positiva. À medida que o pH aumenta, o grupo carboxila perde seu próton primeiro, gerando a forma zwitteriônica, na qual a carga positiva do grupo amino é compensada pela carga negativa do grupo carboxilato. Em pH ainda mais elevado, ocorre a desprotonação do grupo amino, levando a uma carga líquida negativa. Essa transição gradual entre estados de carga é descrita pelas curvas de titulação dos aminoácidos [Figura], que permitem visualizar a relação entre pH e grau de ionização.
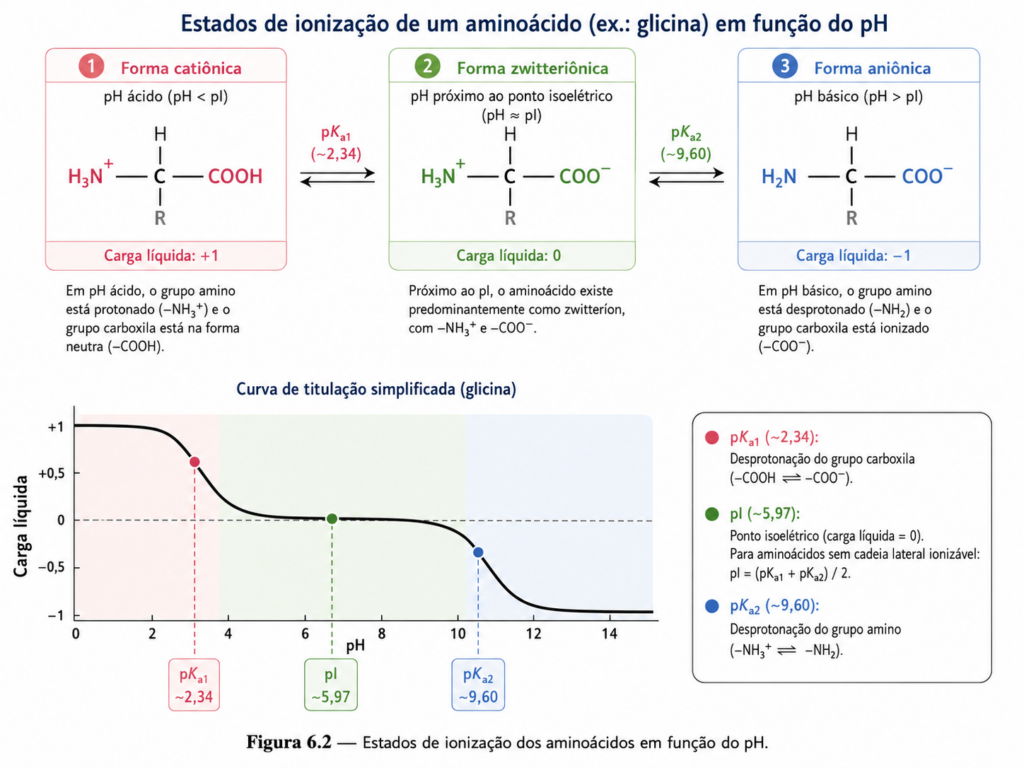
Um conceito particularmente relevante nesse contexto é o ponto isoelétrico (pI), definido como o pH no qual a carga líquida do aminoácido é zero. Nesse ponto, a molécula apresenta mobilidade mínima em um campo elétrico, propriedade amplamente explorada em técnicas laboratoriais como a eletroforese. O valor do pI varia entre os aminoácidos, dependendo da presença de grupos ionizáveis adicionais nas cadeias laterais. Aminoácidos ácidos, como ácido aspártico e ácido glutâmico, apresentam pI mais baixos, enquanto aminoácidos básicos, como lisina e arginina, possuem pI mais elevados.
As cadeias laterais desempenham um papel decisivo na modulação das propriedades químicas dos aminoácidos. Algumas contêm grupos funcionais capazes de participar de reações específicas, como grupos hidroxila (serina e treonina), sulfidrila (cisteína) e anéis aromáticos (fenilalanina, tirosina e triptofano). Esses grupos podem atuar como nucleófilos, participar de reações redox ou formar ligações covalentes, ampliando significativamente a gama de funções químicas disponíveis nas proteínas. Em especial, a cisteína merece destaque por sua capacidade de formar pontes dissulfeto, ligações covalentes que estabilizam estruturas proteicas em ambientes extracelulares [Molécula].
Outro aspecto crítico é a influência do microambiente sobre os valores de pKa. Em solução aquosa ideal, os valores de pKa são relativamente constantes; entretanto, no interior de proteínas, esses valores podem sofrer deslocamentos significativos devido a interações com outros grupos próximos, à polaridade local e à exposição ao solvente. Esse fenômeno é essencial para a atividade catalítica de muitas enzimas, nas quais resíduos específicos atuam como doadores ou aceitadores de prótons em condições finamente ajustadas.
Além das propriedades ácido-base, os aminoácidos também exibem comportamentos distintos em termos de polaridade e capacidade de interação com o solvente. Aminoácidos hidrofóbicos tendem a evitar o contato com a água, favorecendo interações entre si, enquanto aminoácidos hidrofílicos estabelecem ligações de hidrogênio e interações eletrostáticas com o meio aquoso. Essa dualidade é um dos princípios fundamentais que governam o dobramento proteico e a formação de estruturas tridimensionais estáveis.
Do ponto de vista funcional, essas propriedades químicas não atuam isoladamente, mas de forma integrada. A combinação de estados de ionização, características das cadeias laterais e interações com o ambiente define o comportamento coletivo dos aminoácidos quando incorporados em proteínas. Em sistemas biológicos, isso se traduz na capacidade de proteínas responderem a variações de pH, interagirem seletivamente com outras moléculas e desempenharem funções catalíticas altamente específicas.
Portanto, as propriedades químicas dos aminoácidos constituem a base para a compreensão de fenômenos mais complexos da bioquímica, como a estabilidade estrutural das proteínas, a catálise enzimática e a regulação metabólica. Ignorar essa base compromete a interpretação de praticamente todos os processos moleculares subsequentes, o que torna esse tópico um dos pilares conceituais da disciplina.
6.3 Ligações peptídicas e formação de polipeptídeos #
A transição entre a química dos aminoácidos isolados e a complexidade estrutural das proteínas ocorre por meio de um evento fundamental: a formação da ligação peptídica. Essa ligação covalente estabelece a conexão entre aminoácidos, permitindo a construção de cadeias lineares — os polipeptídeos — que servirão de base para todos os níveis superiores de organização proteica. Embora, à primeira vista, esse processo pareça apenas uma condensação simples entre dois grupos funcionais, sua natureza química revela implicações estruturais profundas.
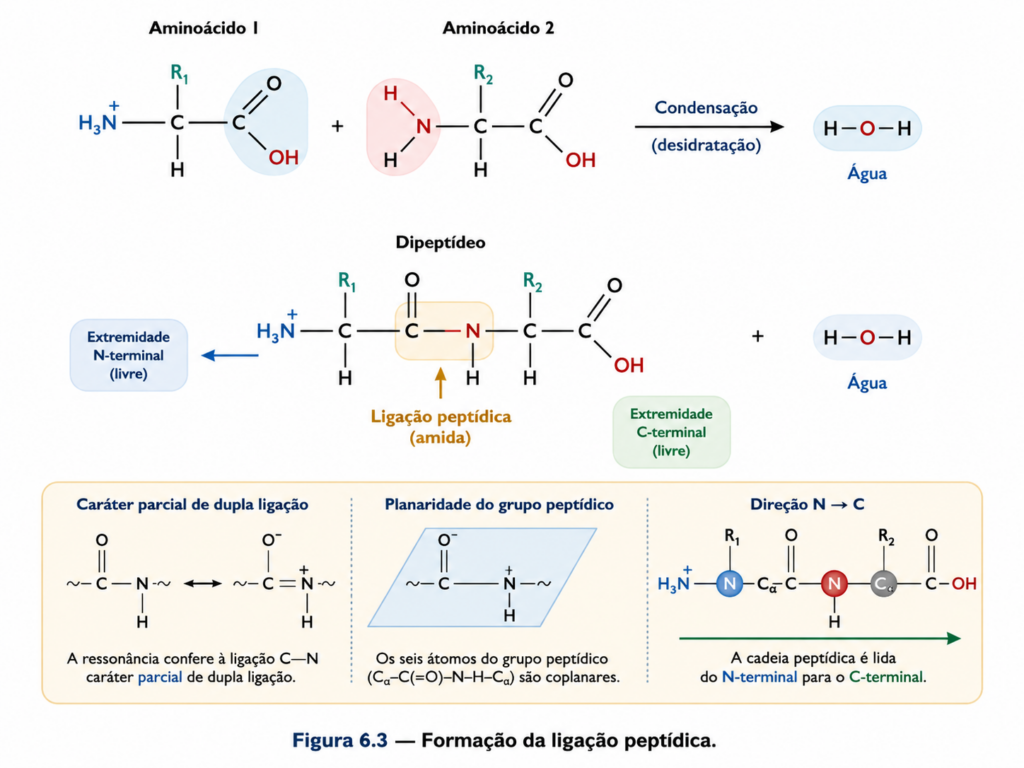
A ligação peptídica forma-se por uma reação de condensação entre o grupo carboxila de um aminoácido e o grupo amino de outro, resultando na liberação de uma molécula de água e na criação de uma ligação amida (–CO–NH–) [Equação]. Esse processo, energeticamente desfavorável em condições isoladas, é catalisado no ambiente celular por maquinarias altamente especializadas, como o ribossomo, que acoplam a síntese proteica ao gasto de energia metabólica. O resultado é uma cadeia polipeptídica com direção definida, que se estende do terminal amino (N-terminal) ao terminal carboxila (C-terminal), uma convenção essencial para a descrição e interpretação das sequências proteicas.
Do ponto de vista eletrônico, a ligação peptídica apresenta uma característica crítica: o compartilhamento de elétrons entre o oxigênio carbonílico e o nitrogênio amídico gera um sistema de ressonância que confere à ligação um caráter parcial de dupla ligação. Essa propriedade restringe a rotação livre em torno do eixo C–N, tornando o grupo peptídico planar. Como consequência, os átomos envolvidos — carbono carbonílico, oxigênio, nitrogênio e hidrogênio — situam-se aproximadamente no mesmo plano, formando uma unidade estrutural rígida [Figura]. Essa rigidez é um dos principais determinantes da arquitetura tridimensional das proteínas.
Embora a rotação em torno da ligação peptídica seja limitada, existe liberdade conformacional em torno das ligações adjacentes ao carbono α, descritas pelos ângulos diedros φ (phi) e ψ (psi). Esses ângulos definem a orientação relativa entre os planos peptídicos consecutivos e são responsáveis pela diversidade conformacional das cadeias polipeptídicas. No entanto, nem todas as combinações desses ângulos são energeticamente favoráveis, devido a impedimentos estéricos entre os átomos. Essa restrição conformacional é classicamente representada no diagrama de Ramachandran, que delimita as regiões permitidas para os ângulos φ e ψ, estabelecendo a base geométrica para a formação de estruturas secundárias como hélices α e folhas β.
Outro aspecto relevante da ligação peptídica é sua polaridade intrínseca. O grupo carbonila apresenta caráter parcialmente negativo, enquanto o grupo amida possui caráter parcialmente positivo, criando um dipolo ao longo da ligação. Essa polaridade permite a formação de ligações de hidrogênio entre diferentes segmentos da cadeia polipeptídica, interações que são fundamentais para a estabilização das estruturas secundárias das proteínas. Assim, a própria natureza da ligação peptídica já contém os elementos necessários para a organização estrutural em níveis mais complexos.
A sequência linear de aminoácidos em um polipeptídeo — determinada geneticamente — define a chamada estrutura primária da proteína. No entanto, essa sequência não é apenas uma lista de resíduos; ela representa um código químico que contém todas as informações necessárias para o dobramento e a função da proteína. A disposição específica de aminoácidos com diferentes propriedades — hidrofóbicas, polares, carregadas — orienta a cadeia durante o processo de enovelamento, conduzindo-a a uma conformação tridimensional energeticamente estável.
Além disso, é importante considerar que as cadeias polipeptídicas não existem isoladamente em condições fisiológicas. Durante e após sua síntese, elas interagem com o ambiente celular, com outras proteínas e, frequentemente, com cofatores ou íons metálicos. Essas interações começam a influenciar a estrutura ainda no nível da cadeia linear, mostrando que a formação da ligação peptídica é apenas o primeiro passo em um processo altamente coordenado de construção molecular.
Portanto, a ligação peptídica não deve ser interpretada apenas como um elo químico entre aminoácidos, mas como um elemento estruturante que impõe restrições geométricas, estabelece direcionalidade e cria as condições necessárias para a emergência da complexidade proteica. A partir dessa base, a cadeia polipeptídica adquire potencial para se organizar em estruturas hierárquicas que, em última instância, sustentam praticamente todas as funções biológicas conhecidas.
6.4 Estrutura primária das proteínas #
A estrutura primária de uma proteína corresponde à sequência linear específica de aminoácidos que compõem sua cadeia polipeptídica. Embora essa definição pareça simples, ela representa, na prática, o nível mais fundamental de organização proteica e contém toda a informação necessária para determinar os níveis estruturais superiores. Em outras palavras, a sequência de aminoácidos não é apenas uma ordem arbitrária de resíduos, mas um código químico altamente organizado, resultado direto da informação genética codificada no DNA e expressa por meio dos processos de transcrição e tradução.
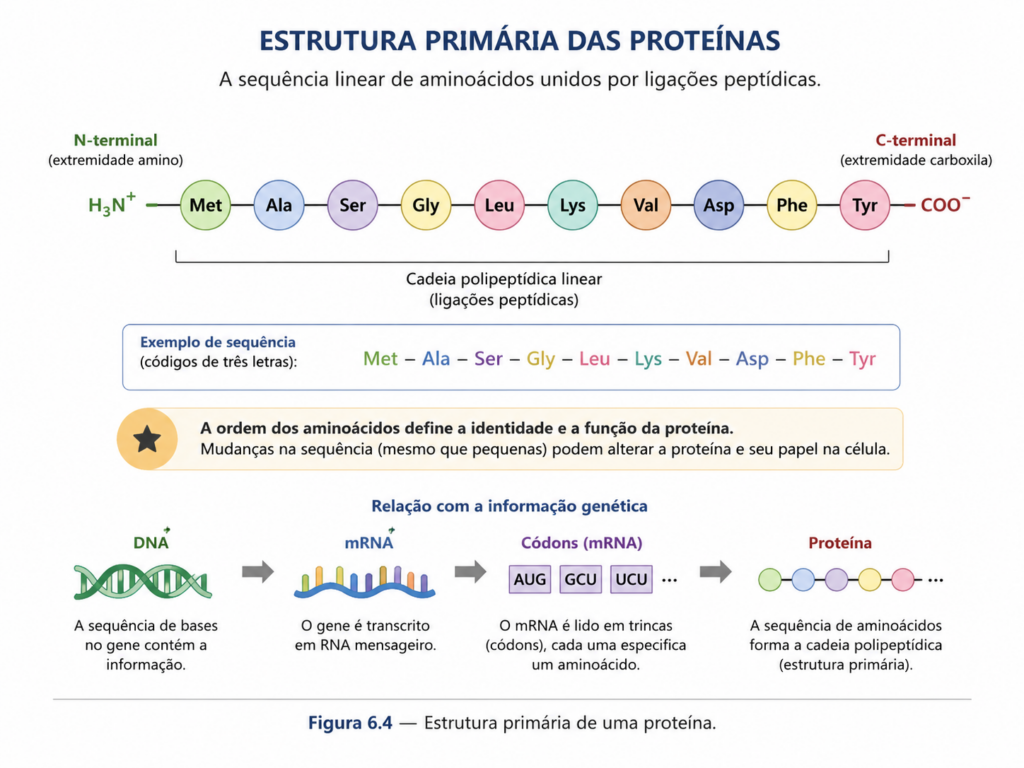
Essa sequência é estabelecida durante a síntese proteica no ribossomo, onde cada aminoácido é incorporado de acordo com a sequência de códons do RNA mensageiro, seguindo o código genético universal . A precisão desse processo é essencial, pois mesmo alterações pontuais — como a substituição de um único aminoácido — podem provocar mudanças significativas na estrutura e na função da proteína. Esse fenômeno é observado em diversas condições patológicas, nas quais mutações gênicas resultam em proteínas estruturalmente alteradas e funcionalmente comprometidas.
A importância da estrutura primária está diretamente relacionada à natureza das cadeias laterais dos aminoácidos que a compõem. A distribuição de resíduos hidrofóbicos, polares e carregados ao longo da cadeia influencia o comportamento da proteína em solução aquosa e direciona seu processo de dobramento. Aminoácidos hidrofóbicos tendem a se agrupar no interior da proteína, afastando-se do meio aquoso, enquanto resíduos polares e carregados permanecem expostos ao solvente. Esse padrão de distribuição não é aleatório, mas sim resultado de pressões evolutivas que selecionaram sequências capazes de formar estruturas estáveis e funcionalmente eficientes.
Além disso, a estrutura primária também determina a presença de motivos estruturais e domínios funcionais. Certas sequências recorrentes de aminoácidos estão associadas a funções específicas, como ligação a ligantes, atividade catalítica ou interação com outras proteínas. Esses padrões, conhecidos como motivos conservados, são frequentemente identificados em proteínas de diferentes organismos, indicando sua importância funcional e sua conservação ao longo da evolução. A análise comparativa de sequências permite, portanto, inferir relações evolutivas e prever funções de proteínas ainda não caracterizadas.
Outro aspecto relevante é que a estrutura primária define o posicionamento de resíduos críticos para interações químicas específicas. Aminoácidos como cisteína podem formar pontes dissulfeto, contribuindo para a estabilidade estrutural, enquanto resíduos como histidina, ácido aspártico e serina frequentemente participam de mecanismos catalíticos em enzimas. A posição relativa desses resíduos na sequência linear é determinante para que, após o dobramento, eles se encontrem no espaço tridimensional de forma adequada para exercer sua função.
Do ponto de vista experimental, a determinação da estrutura primária foi um marco na história da bioquímica. Métodos clássicos, como a degradação de Edman, permitiram sequenciar proteínas pela remoção sequencial de resíduos do terminal amino, enquanto técnicas modernas, como espectrometria de massas, possibilitam análises rápidas e altamente precisas de sequências proteicas . Essas abordagens não apenas ampliaram o conhecimento sobre proteínas individuais, mas também viabilizaram a construção de grandes bancos de dados de sequências, fundamentais para estudos de bioinformática e biologia evolutiva.
A relação entre estrutura primária e função foi demonstrada de forma inequívoca em experimentos clássicos de dobramento proteico, nos quais proteínas desnaturadas foram capazes de recuperar sua conformação nativa apenas com base em sua sequência de aminoácidos. Esse princípio, frequentemente associado ao trabalho de Anfinsen, estabelece que a estrutura tridimensional de uma proteína é determinada intrinsecamente por sua sequência primária, desde que as condições ambientais sejam adequadas.
Entretanto, é importante reconhecer que, em sistemas biológicos reais, o dobramento proteico pode ser assistido por chaperonas moleculares e influenciado por fatores celulares, o que adiciona camadas de complexidade ao processo. Ainda assim, a sequência de aminoácidos permanece como o determinante central da estrutura e da função proteica.
Portanto, a estrutura primária deve ser entendida como o nível organizacional que conecta diretamente a informação genética à função biológica. Ela estabelece as bases para todas as interações e transformações subsequentes que a proteína sofrerá, sendo, ao mesmo tempo, o ponto de partida e o elemento definidor de sua identidade molecular. Qualquer análise estrutural ou funcional de proteínas que desconsidere a importância da sequência primária estará, inevitavelmente, comprometida em sua profundidade e precisão.
6.5 Estrutura secundária #
A estrutura secundária das proteínas representa o primeiro nível de organização tridimensional que emerge a partir da sequência linear de aminoácidos. Diferentemente da estrutura primária, que descreve apenas a ordem dos resíduos, a estrutura secundária refere-se a padrões locais de dobramento da cadeia polipeptídica, estabilizados principalmente por ligações de hidrogênio entre os grupos da espinha dorsal peptídica. Esses padrões não dependem diretamente das cadeias laterais, mas são fortemente influenciados por elas, na medida em que impõem restrições estéricas e eletrônicas ao arranjo espacial da cadeia.
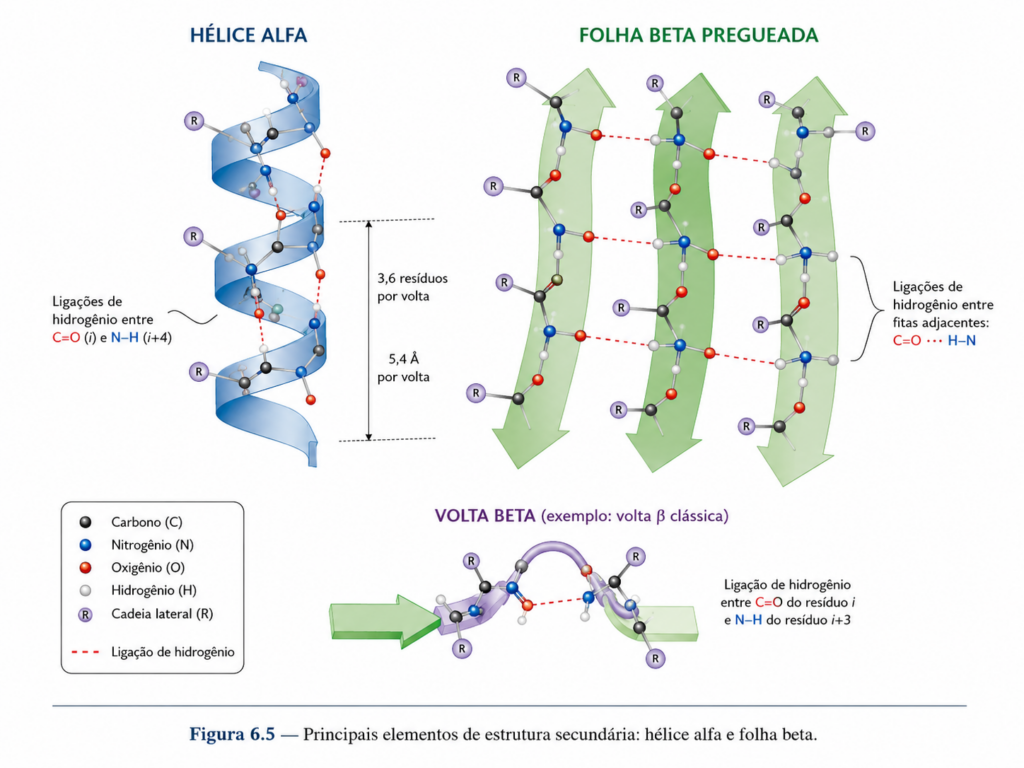
O fundamento químico da estrutura secundária está na própria natureza da ligação peptídica. Como discutido anteriormente, essa ligação possui caráter parcial de dupla ligação, o que confere rigidez e planaridade ao grupo peptídico. Essa restrição limita as possibilidades conformacionais da cadeia e favorece a formação de arranjos regulares e repetitivos. Ao longo da cadeia polipeptídica, os grupos carbonila (C=O) e amida (N–H) tornam-se capazes de estabelecer ligações de hidrogênio entre si, criando redes estabilizadoras que sustentam essas estruturas locais.
Entre os principais tipos de estrutura secundária, destacam-se a hélice α e a folha β, que constituem os motivos estruturais mais recorrentes nas proteínas. A hélice α é uma estrutura helicoidal na qual a cadeia polipeptídica se enrola sobre si mesma, formando uma espiral compacta. Nesse arranjo, cada grupo carbonila estabelece uma ligação de hidrogênio com o grupo amida de um aminoácido localizado quatro resíduos à frente na sequência. Esse padrão regular gera uma estrutura altamente estável, com as cadeias laterais projetando-se para fora da hélice, minimizando interferências estéricas [Figura]. A hélice α é particularmente favorecida por aminoácidos com cadeias laterais pequenas e não ramificadas, enquanto resíduos volumosos ou com restrições conformacionais, como a prolina, tendem a desestabilizá-la.
As folhas β, por sua vez, consistem em segmentos de cadeia polipeptídica estendidos que se alinham lado a lado, formando uma estrutura semelhante a uma lâmina pregueada. Esses segmentos, denominados fitas β, podem estar orientados em paralelo ou antiparalelo, dependendo da direção das cadeias na sequência. As ligações de hidrogênio ocorrem entre grupos peptídicos de fitas adjacentes, conferindo estabilidade ao conjunto. Diferentemente da hélice α, as cadeias laterais nas folhas β alternam-se acima e abaixo do plano da estrutura, o que permite a formação de interfaces de interação com outras regiões da proteína ou com o meio externo.
Além desses dois motivos principais, existem elementos estruturais adicionais, como voltas β e regiões desordenadas, que desempenham papéis importantes na organização global da proteína. As voltas β são segmentos curtos que promovem mudanças bruscas na direção da cadeia polipeptídica, permitindo a compactação da estrutura. Frequentemente, envolvem aminoácidos como glicina e prolina, que favorecem essas mudanças conformacionais. Já as regiões desordenadas, embora não apresentem um padrão estrutural fixo, são funcionalmente relevantes, participando de interações dinâmicas e processos regulatórios.
A formação da estrutura secundária não ocorre de maneira isolada, mas sim como parte de um processo contínuo de dobramento da proteína. Esses elementos estruturais surgem rapidamente durante o enovelamento inicial da cadeia e servem como blocos fundamentais para a construção da estrutura terciária. A estabilidade dessas estruturas depende não apenas das ligações de hidrogênio, mas também de fatores como o ambiente químico, a presença de solvente e as interações com outras partes da molécula.
É importante destacar que a estrutura secundária reflete um equilíbrio entre restrições geométricas e interações químicas. Nem todas as sequências de aminoácidos são capazes de formar hélices α ou folhas β de maneira eficiente; a propensão para cada tipo de estrutura depende da composição e da ordem dos resíduos. Esse comportamento pode ser previsto, em certa medida, por análises computacionais e estudos experimentais, sendo uma ferramenta importante na bioinformática estrutural.
Do ponto de vista funcional, a estrutura secundária contribui diretamente para a estabilidade mecânica e a especificidade das proteínas. Em proteínas fibrosas, como o colágeno, padrões repetitivos de estrutura secundária conferem resistência e elasticidade. Em proteínas globulares, esses elementos estruturais organizam-se de maneira compacta, formando domínios funcionais capazes de realizar atividades biológicas específicas.
Portanto, a estrutura secundária deve ser entendida como um nível intermediário essencial entre a sequência linear e a conformação tridimensional completa das proteínas. Ela traduz as propriedades químicas da ligação peptídica em padrões estruturais definidos, estabelecendo as bases para a complexidade funcional que será plenamente desenvolvida nos níveis estruturais subsequentes.
6.6 Estrutura terciária #
A estrutura terciária corresponde ao arranjo tridimensional completo de uma cadeia polipeptídica, resultante do dobramento global que organiza os elementos de estrutura secundária em uma conformação específica e funcional. Se a estrutura primária representa o código e a estrutura secundária os padrões locais, a estrutura terciária é a materialização espacial desse código, onde a proteína adquire sua identidade funcional. Trata-se, portanto, do nível em que a bioquímica estrutural se conecta diretamente à atividade biológica.
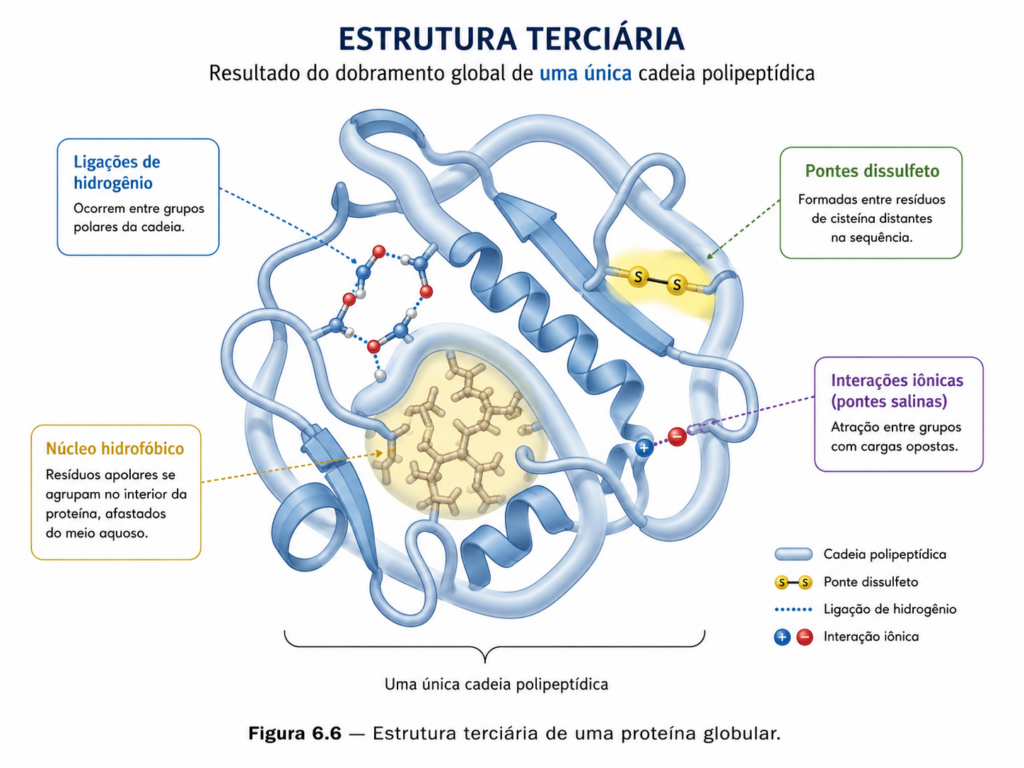
Esse dobramento não ocorre de forma aleatória, mas é guiado por um princípio termodinâmico fundamental: a proteína tende a assumir a conformação de menor energia livre possível no ambiente em que se encontra. Durante esse processo, a cadeia polipeptídica explora múltiplas conformações até atingir um estado estável, frequentemente descrito como um mínimo energético em uma “paisagem de enovelamento” [Figura]. A sequência de aminoácidos determina o percurso dessa trajetória, direcionando interações que conduzem ao estado nativo.
As forças que estabilizam a estrutura terciária são diversas e atuam de maneira cooperativa. Entre elas, destacam-se as interações hidrofóbicas, que desempenham papel central no dobramento de proteínas em meio aquoso. Aminoácidos com cadeias laterais apolares tendem a se agrupar no interior da proteína, afastando-se da água, formando um núcleo hidrofóbico compacto. Esse efeito não é simplesmente uma atração entre resíduos apolares, mas uma consequência da reorganização da água ao redor de superfícies hidrofóbicas, o que torna energeticamente favorável o seu agrupamento.
Além das interações hidrofóbicas, as ligações de hidrogênio contribuem para a estabilização da estrutura terciária, tanto entre grupos da cadeia principal quanto entre cadeias laterais. Interações eletrostáticas, como pontes salinas entre resíduos carregados positivamente e negativamente, também são relevantes, especialmente em regiões expostas ao solvente. Em ambientes menos polares, como o interior da proteína, essas interações podem se tornar ainda mais fortes, influenciando significativamente a conformação final.
Outro tipo de interação importante é a formação de pontes dissulfeto, ligações covalentes entre resíduos de cisteína que atuam como “travas” estruturais, conferindo estabilidade adicional, especialmente em proteínas extracelulares. Essas ligações são particularmente relevantes em condições onde a proteína está sujeita a variações de pH, temperatura ou agentes desnaturantes, funcionando como elementos de resistência estrutural.
A estrutura terciária também se organiza frequentemente em unidades funcionais denominadas domínios. Um domínio é uma região da proteína que se dobra de forma relativamente independente e que, muitas vezes, está associada a uma função específica, como ligação a ligantes ou atividade catalítica. Proteínas maiores podem conter múltiplos domínios, permitindo a combinação de diferentes funções em uma única molécula. Essa modularidade estrutural é um dos fatores que contribuem para a diversidade funcional das proteínas ao longo da evolução.
A determinação experimental da estrutura terciária foi um dos grandes avanços da bioquímica moderna. Técnicas como cristalografia de raios X, ressonância magnética nuclear (RMN) e, mais recentemente, criomicroscopia eletrônica, permitiram visualizar proteínas em nível atômico, revelando detalhes das interações que sustentam sua conformação . Esses métodos não apenas confirmaram princípios teóricos do dobramento proteico, mas também abriram caminho para aplicações em áreas como design de fármacos e engenharia de proteínas.
É importante reconhecer que a estrutura terciária não é rigidamente fixa. Muitas proteínas exibem flexibilidade conformacional, podendo alternar entre diferentes estados estruturais em resposta à ligação de ligantes ou mudanças no ambiente. Essa dinâmica é essencial para funções como catálise enzimática, transporte molecular e sinalização celular. Em vez de estruturas estáticas, as proteínas devem ser entendidas como sistemas dinâmicos, cuja função depende da capacidade de transitar entre diferentes conformações.
Além disso, o ambiente celular exerce influência significativa sobre a formação da estrutura terciária. Em muitos casos, proteínas recém-sintetizadas necessitam da ação de chaperonas moleculares, que auxiliam no dobramento correto e evitam agregação indevida. Falhas nesse processo podem levar à formação de estruturas incorretas, frequentemente associadas a doenças relacionadas ao mau enovelamento proteico.
Portanto, a estrutura terciária representa o ponto de convergência entre sequência, interações químicas e ambiente celular. É nesse nível que a proteína adquire sua forma funcional e sua capacidade de interagir de maneira específica com outras moléculas. Qualquer alteração significativa nessa organização — seja por mutação, mudança de pH ou ação de agentes externos — pode comprometer a função da proteína, evidenciando a importância crítica desse nível estrutural na biologia molecular.
6.7 Estrutura quaternária #
A estrutura quaternária representa um nível adicional de organização proteica que emerge quando duas ou mais cadeias polipeptídicas — denominadas subunidades — se associam para formar um complexo funcional. Diferentemente da estrutura terciária, que descreve o dobramento de uma única cadeia, a estrutura quaternária envolve a interação entre múltiplas cadeias já dobradas, resultando em arquiteturas supramoleculares com propriedades que não podem ser atribuídas a uma subunidade isolada. Trata-se, portanto, de um nível de organização onde a cooperação molecular se traduz diretamente em função biológica.
Essas subunidades podem ser idênticas, formando complexos homoméricos, ou distintas, originando estruturas heteroméricas. A associação entre elas é mediada por um conjunto de interações não covalentes semelhantes às que estabilizam a estrutura terciária: interações hidrofóbicas, ligações de hidrogênio e interações eletrostáticas. Em alguns casos, ligações covalentes, como pontes dissulfeto intercadeias, também contribuem para a estabilidade do complexo. A interface entre subunidades é altamente específica, refletindo um encaixe estrutural preciso que garante a formação do complexo correto.
Um dos aspectos mais relevantes da estrutura quaternária é a possibilidade de surgimento de propriedades emergentes, isto é, características funcionais que não estão presentes em subunidades isoladas. O exemplo clássico é a hemoglobina, uma proteína tetramérica responsável pelo transporte de oxigênio no sangue. Cada subunidade da hemoglobina é capaz de ligar oxigênio individualmente; no entanto, a associação entre elas permite um comportamento cooperativo, no qual a ligação de uma molécula de oxigênio facilita a ligação das demais. Esse fenômeno, conhecido como cooperatividade, resulta de mudanças conformacionais transmitidas entre as subunidades, alterando suas afinidades de ligação [Figura].
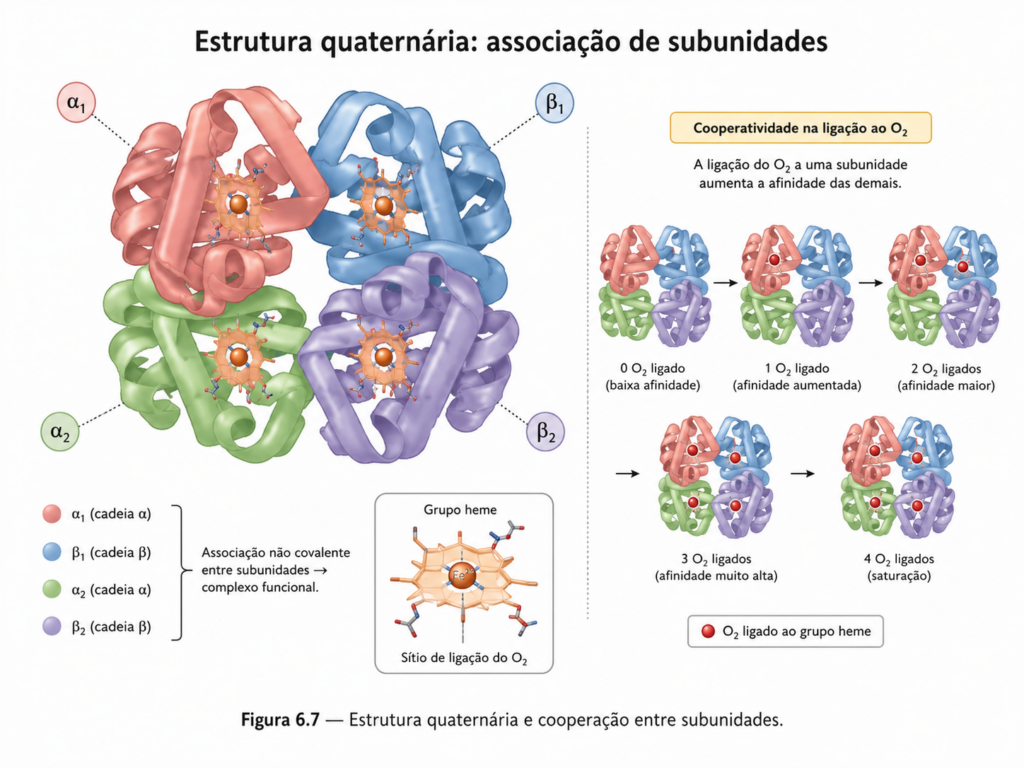
Essa capacidade de transmitir alterações estruturais entre diferentes partes do complexo está relacionada ao conceito de alosteria. Em proteínas oligoméricas, a ligação de um ligante em um sítio específico pode induzir mudanças conformacionais que afetam sítios distantes, modulando a atividade da proteína. Esse mecanismo é amplamente utilizado na regulação metabólica, permitindo respostas rápidas e coordenadas a variações no ambiente celular.
A organização quaternária também apresenta padrões de simetria que refletem princípios de eficiência estrutural. Muitos complexos proteicos exibem simetria rotacional ou repetitiva, o que reduz a complexidade genética necessária para sua codificação e favorece a estabilidade estrutural. Essa simetria não é apenas estética, mas funcional, contribuindo para a formação de estruturas robustas e energeticamente favoráveis.
Do ponto de vista funcional, a formação de estruturas quaternárias oferece vantagens significativas. A associação de múltiplas subunidades pode aumentar a estabilidade da proteína, proteger regiões sensíveis contra degradação e permitir a integração de diferentes funções em um único complexo. Além disso, a modularidade associada à estrutura quaternária facilita a evolução de novas funções, uma vez que subunidades podem ser recombinadas ou modificadas sem comprometer completamente a integridade estrutural do sistema.
A dinâmica dessas associações também é um aspecto relevante. Em muitos casos, as interações entre subunidades são reversíveis, permitindo que complexos se formem e se dissociem conforme as necessidades celulares. Esse comportamento é fundamental em processos como sinalização celular, montagem de complexos enzimáticos e regulação de vias metabólicas. Assim, a estrutura quaternária não deve ser vista como um estado fixo, mas como um equilíbrio dinâmico entre associação e dissociação.
Do ponto de vista experimental, a caracterização da estrutura quaternária exige abordagens que permitam observar a proteína em seu estado funcional completo. Técnicas como cristalografia de raios X, criomicroscopia eletrônica e métodos de análise de massa e interação molecular têm sido fundamentais para elucidar a organização de complexos proteicos . Esses estudos revelaram a complexidade e a diversidade das arquiteturas quaternárias, desde pequenos dímeros até grandes máquinas moleculares compostas por dezenas de subunidades.
Portanto, a estrutura quaternária representa um nível de integração estrutural onde a função biológica se torna, em muitos casos, inseparável da cooperação entre múltiplas cadeias polipeptídicas. É nesse nível que emergem propriedades como cooperatividade, alosteria e regulação fina da atividade proteica, consolidando a ideia de que, na bioquímica, o todo frequentemente é mais do que a soma de suas partes.
6.8 Dobramento de proteínas (protein folding) #
O dobramento de proteínas é o processo pelo qual uma cadeia polipeptídica recém-sintetizada adquire sua conformação tridimensional funcional. Esse fenômeno representa uma das transições mais críticas da bioquímica molecular, pois conecta diretamente a informação contida na sequência de aminoácidos à estrutura e, consequentemente, à função biológica. Embora a síntese da cadeia ocorra de forma linear no ribossomo, sua funcionalidade depende da aquisição de uma estrutura espacial específica, que emerge de um processo altamente coordenado e, em muitos aspectos, ainda não completamente compreendido.
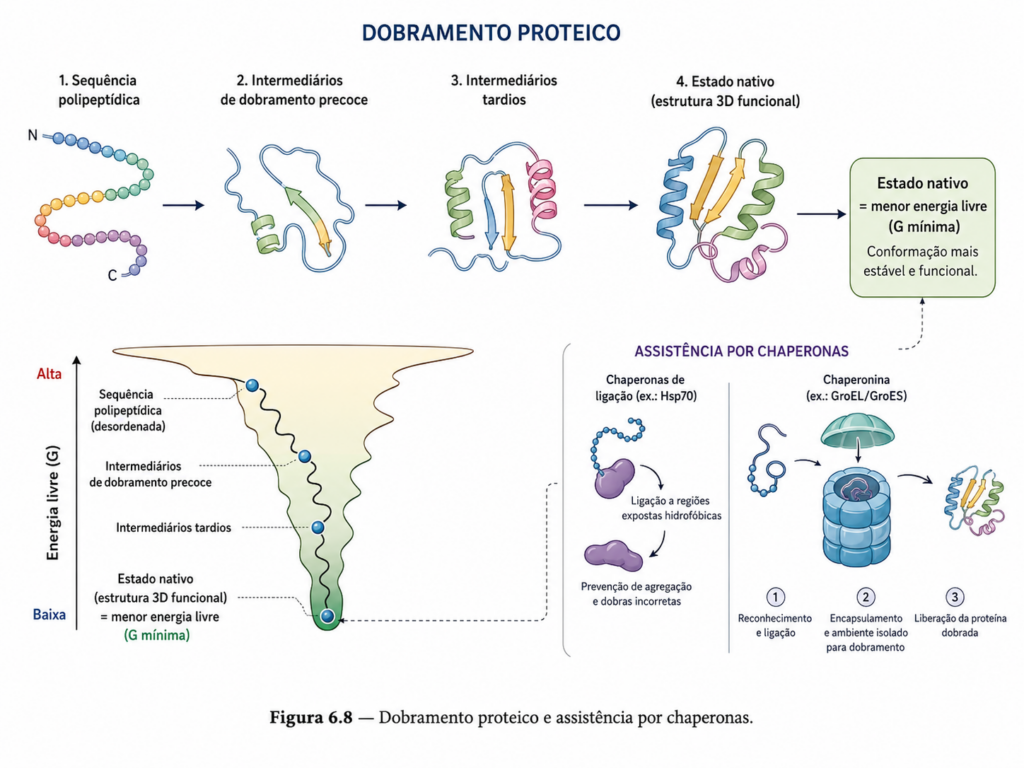
Do ponto de vista termodinâmico, o dobramento é guiado pela busca de um estado de menor energia livre. A proteína explora um grande número de conformações possíveis até alcançar uma estrutura estável, denominada estado nativo. Esse processo é frequentemente descrito por meio do conceito de “paisagem energética”, na qual a proteína percorre um funil conformacional que a conduz de estados altamente desordenados para uma conformação mais organizada e energeticamente favorável [Figura]. Esse modelo resolve, em parte, o chamado paradoxo de Levinthal, que argumenta que uma busca aleatória por todas as conformações possíveis levaria um tempo incompatível com a realidade biológica. Na prática, o dobramento ocorre de maneira dirigida, com trajetórias preferenciais determinadas pela própria sequência de aminoácidos.
Durante o processo de enovelamento, estruturas secundárias como hélices α e folhas β formam-se rapidamente, servindo como núcleos iniciais de organização. A partir desses elementos, interações de longo alcance entre diferentes regiões da cadeia promovem o colapso hidrofóbico, no qual resíduos apolares se agrupam no interior da proteína, enquanto grupos polares permanecem expostos ao solvente. Esse rearranjo é um dos principais motores do dobramento, sendo complementado por interações eletrostáticas, ligações de hidrogênio e, em alguns casos, pela formação de pontes dissulfeto.
Entretanto, o ambiente celular impõe desafios adicionais ao dobramento proteico. Em um meio altamente concentrado e dinâmico, cadeias polipeptídicas recém-sintetizadas estão sujeitas ao risco de agregação indevida, especialmente quando regiões hidrofóbicas ficam temporariamente expostas. Para evitar esse problema, as células utilizam proteínas auxiliares conhecidas como chaperonas moleculares. Essas moléculas não fazem parte da estrutura final da proteína, mas atuam facilitando o dobramento correto, impedindo interações inadequadas e, em alguns casos, fornecendo um ambiente protegido para o enovelamento.
Entre as chaperonas mais estudadas estão as chaperoninas, complexos proteicos que formam estruturas semelhantes a “câmaras” onde a proteína pode se dobrar isoladamente do meio externo. Esses sistemas utilizam energia derivada da hidrólise de ATP para promover ciclos de ligação e liberação da proteína em dobramento, aumentando a eficiência e a fidelidade do processo. A presença dessas máquinas moleculares evidencia que, embora a sequência de aminoácidos contenha a informação necessária para o dobramento, o contexto celular é determinante para sua execução adequada.
O dobramento proteico também pode envolver estados intermediários, nos quais a proteína apresenta organização parcial. Esses intermediários são transitórios, mas podem desempenhar papel importante na trajetória de enovelamento. Em condições normais, a proteína progride desses estados para o estado nativo; entretanto, falhas nesse processo podem levar à formação de estruturas mal dobradas. Essas formas aberrantes tendem a se agregar, formando complexos insolúveis que estão associados a diversas doenças, como enfermidades neurodegenerativas e distúrbios por proteínas príonicas.
Outro ponto crítico é que o estado nativo de uma proteína nem sempre corresponde a uma estrutura rígida única. Muitas proteínas apresentam flexibilidade conformacional e podem existir em múltiplos estados estruturais relacionados, alternando entre eles conforme as condições do ambiente ou a presença de ligantes. Essa dinâmica é essencial para funções como catálise enzimática, sinalização celular e interação com outras macromoléculas.
Do ponto de vista experimental e computacional, o estudo do dobramento proteico avançou significativamente nas últimas décadas. Métodos como espectroscopia, calorimetria e simulações moleculares permitiram caracterizar etapas do processo e identificar fatores que influenciam a estabilidade estrutural. Mais recentemente, abordagens baseadas em inteligência artificial têm permitido prever estruturas tridimensionais a partir da sequência de aminoácidos, evidenciando o grau de determinismo contido na estrutura primária.
Portanto, o dobramento de proteínas deve ser entendido como um processo integrado, no qual propriedades químicas intrínsecas da cadeia polipeptídica interagem com o ambiente celular para produzir estruturas funcionais. Trata-se de um fenômeno que combina princípios de termodinâmica, cinética e interação molecular, sendo essencial para a manutenção da vida em nível molecular. Qualquer falha nesse processo compromete diretamente a funcionalidade proteica, reforçando sua importância central na bioquímica.
6.9 Desnaturação e estabilidade proteica #
A estrutura tridimensional de uma proteína, embora altamente organizada e funcional, não é rigidamente fixa. Ela resulta de um equilíbrio delicado entre forças estabilizadoras e fatores ambientais que podem, sob determinadas condições, levar à sua desorganização. Esse processo de perda da conformação nativa é denominado desnaturação, e sua compreensão é essencial para interpretar tanto a funcionalidade quanto a fragilidade das proteínas em sistemas biológicos.
A desnaturação consiste na ruptura das interações que mantêm a estrutura secundária, terciária e quaternária de uma proteína, sem, em geral, afetar a integridade da estrutura primária. Isso significa que a sequência de aminoácidos permanece intacta, mas a organização espacial da molécula é perdida. Como consequência direta, a proteína deixa de exercer sua função biológica, uma vez que a atividade depende criticamente de sua conformação tridimensional.
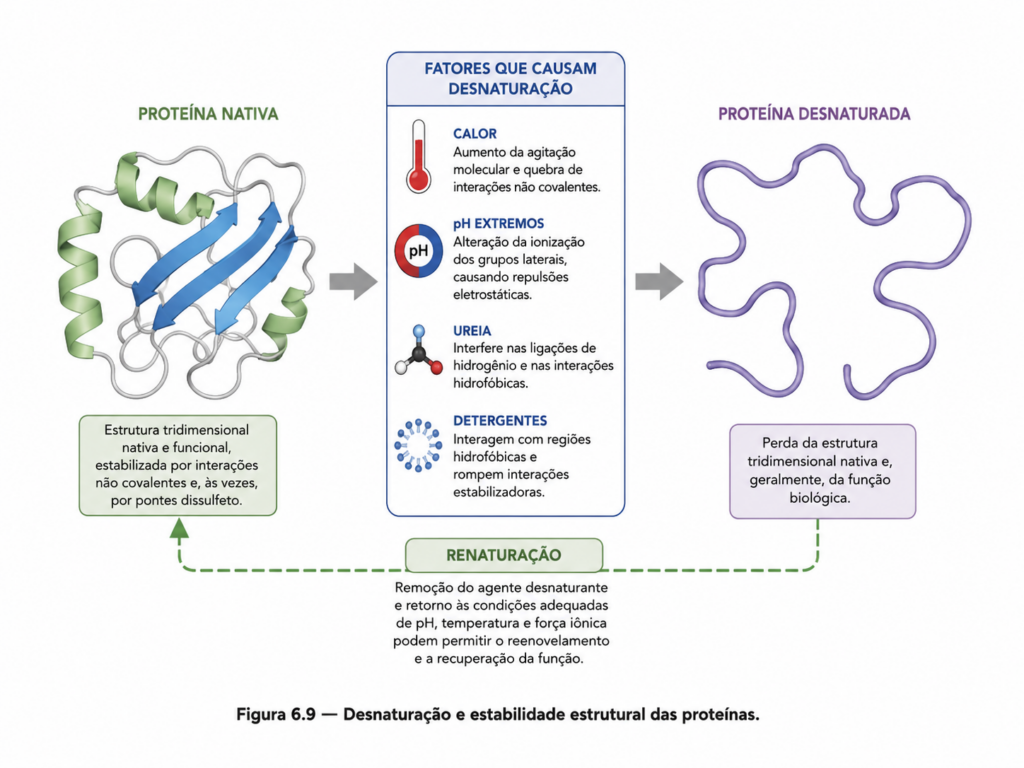
Diversos fatores podem induzir a desnaturação proteica. Entre os mais relevantes estão a temperatura, o pH e a presença de agentes químicos. O aumento da temperatura intensifica o movimento molecular, enfraquecendo interações não covalentes como ligações de hidrogênio e interações hidrofóbicas. Em temperaturas suficientemente elevadas, essas interações são rompidas, levando ao desenovelamento da proteína. Esse fenômeno é facilmente observado em sistemas cotidianos, como a coagulação de proteínas durante o aquecimento de alimentos.
O pH também exerce influência significativa sobre a estabilidade proteica. Alterações no pH modificam o estado de ionização dos grupos funcionais dos aminoácidos, afetando interações eletrostáticas e ligações de hidrogênio. Em valores extremos de pH, essas mudanças podem desestabilizar a estrutura da proteína, promovendo sua desnaturação. Esse efeito é particularmente relevante em sistemas biológicos, nos quais pequenas variações de pH podem ter impacto funcional significativo.
Agentes químicos, como solventes orgânicos, detergentes e compostos caotrópicos (por exemplo, ureia e cloreto de guanidínio), também são capazes de induzir desnaturação. Esses agentes interferem nas interações que estabilizam a estrutura proteica, seja alterando a polaridade do meio, seja competindo por ligações de hidrogênio. Detergentes, em particular, podem se associar a regiões hidrofóbicas da proteína, desorganizando seu núcleo interno e promovendo o desenovelamento.
A estabilidade proteica, por outro lado, refere-se à capacidade da proteína de manter sua estrutura nativa frente a variações ambientais. Essa estabilidade não é absoluta, mas relativa, dependendo do equilíbrio entre forças estabilizadoras e desestabilizadoras. Interações hidrofóbicas, ligações de hidrogênio, interações iônicas e pontes dissulfeto atuam de forma cooperativa para manter a estrutura, enquanto fatores externos podem deslocar esse equilíbrio.
Um aspecto importante da desnaturação é sua reversibilidade. Em alguns casos, quando o agente desnaturante é removido, a proteína pode retornar espontaneamente à sua conformação nativa, recuperando sua função. Esse fenômeno reforça o princípio de que a estrutura tridimensional é determinada pela sequência de aminoácidos. No entanto, nem todas as proteínas apresentam essa capacidade de renaturação. Em muitos casos, o desenovelamento expõe regiões hidrofóbicas que promovem agregação irreversível, impedindo o retorno ao estado funcional.
A agregação proteica é uma consequência crítica da desnaturação, especialmente em ambientes celulares. Proteínas mal dobradas ou parcialmente desnaturadas podem interagir entre si de maneira não específica, formando complexos insolúveis. Esses agregados estão associados a diversas patologias, incluindo doenças neurodegenerativas, nas quais o acúmulo de proteínas mal conformadas compromete a função celular.
Do ponto de vista biotecnológico e industrial, o controle da estabilidade proteica é um desafio central. Enzimas utilizadas em processos industriais, por exemplo, precisam manter sua atividade em condições frequentemente adversas, como altas temperaturas ou presença de solventes. Estratégias como engenharia de proteínas e modificação química são empregadas para aumentar a estabilidade, explorando princípios estruturais que reforçam interações internas ou reduzem a flexibilidade excessiva.
Além disso, a estabilidade proteica é um fator crítico no desenvolvimento de fármacos biológicos. Proteínas terapêuticas, como anticorpos monoclonais, devem ser formuladas de modo a preservar sua estrutura durante armazenamento e administração. Pequenas alterações nas condições de pH, temperatura ou composição do solvente podem comprometer sua eficácia, exigindo um controle rigoroso das condições físico-químicas.
Portanto, a desnaturação e a estabilidade proteica devem ser compreendidas como aspectos complementares de um mesmo fenômeno: o equilíbrio estrutural das proteínas. Enquanto a desnaturação revela a vulnerabilidade dessas macromoléculas frente a perturbações ambientais, a estabilidade reflete a robustez de sua organização molecular. A interação entre esses dois aspectos define, em última instância, a capacidade das proteínas de desempenhar suas funções em sistemas biológicos e tecnológicos.
6.10 Proteínas globulares vs fibrosas #
A diversidade estrutural das proteínas se manifesta de forma particularmente evidente quando se compara dois grandes grupos funcionais: as proteínas globulares e as proteínas fibrosas. Essa distinção não é meramente classificatória, mas reflete diferenças profundas na organização estrutural, nas propriedades físico-químicas e, sobretudo, nas funções desempenhadas por essas macromoléculas nos sistemas biológicos.
As proteínas globulares caracterizam-se por apresentarem uma estrutura tridimensional compacta, aproximadamente esférica, resultante de um dobramento altamente organizado da cadeia polipeptídica. Nesse tipo de proteína, os aminoácidos hidrofóbicos tendem a se concentrar no interior da molécula, formando um núcleo estável, enquanto resíduos polares e carregados permanecem expostos ao meio aquoso. Essa organização confere alta solubilidade em água, o que é essencial para proteínas que atuam em ambientes intracelulares ou circulam no plasma sanguíneo.
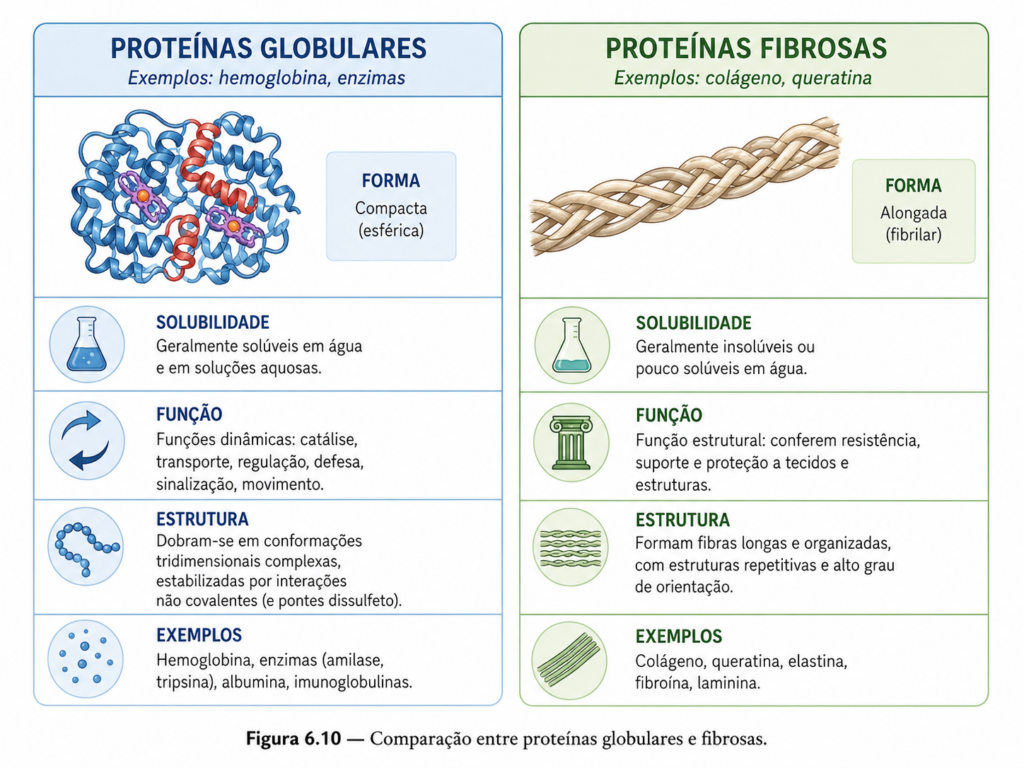
Funcionalmente, as proteínas globulares são extremamente versáteis. Incluem enzimas, proteínas de transporte, anticorpos e proteínas reguladoras. Sua estrutura compacta permite a formação de sítios ativos específicos, capazes de reconhecer e interagir seletivamente com outras moléculas. Essa especificidade estrutural é a base da catálise enzimática e de processos como reconhecimento molecular e sinalização celular. Além disso, muitas proteínas globulares apresentam dinâmica conformacional, podendo alterar sua estrutura em resposta à ligação de ligantes, o que amplia ainda mais seu repertório funcional.
Em contraste, as proteínas fibrosas apresentam uma organização estrutural mais simples e repetitiva, caracterizada por cadeias polipeptídicas alongadas que formam estruturas lineares ou em feixe. Essas proteínas são dominadas por elementos de estrutura secundária repetitivos, como hélices α ou folhas β, organizados de maneira a conferir resistência mecânica e estabilidade estrutural. Diferentemente das proteínas globulares, as proteínas fibrosas possuem baixa solubilidade em água, uma vez que sua função não está associada à mobilidade em solução, mas à formação de estruturas estáveis e duráveis.
As proteínas fibrosas desempenham principalmente funções estruturais. Exemplos clássicos incluem o colágeno, que forma a matriz extracelular e confere resistência a tecidos conjuntivos; a queratina, presente em cabelos, unhas e epiderme; e a fibroína, componente da seda. No caso do colágeno, por exemplo, três cadeias polipeptídicas formam uma hélice tripla altamente organizada, estabilizada por interações específicas e modificações pós-traducionais. Essa estrutura resulta em uma fibra extremamente resistente à tração, adequada à sua função biológica.
A diferença entre proteínas globulares e fibrosas também se reflete em suas propriedades mecânicas e estabilidade. Proteínas globulares são geralmente mais sensíveis a variações de pH e temperatura, podendo desnaturar com relativa facilidade. Já proteínas fibrosas tendem a ser mais resistentes, mantendo sua integridade estrutural mesmo em condições adversas. Essa robustez está diretamente relacionada à repetição estrutural e à presença de interações estabilizadoras extensas ao longo da cadeia.
Do ponto de vista evolutivo, essa dualidade estrutural representa uma adaptação funcional eficiente. Enquanto proteínas globulares evoluíram para desempenhar funções dinâmicas e específicas, exigindo flexibilidade e interação com diversas moléculas, proteínas fibrosas foram selecionadas para fornecer suporte estrutural e resistência mecânica, onde a estabilidade é mais crítica do que a flexibilidade.
É importante destacar que essa classificação, embora útil, não é absolutamente rígida. Algumas proteínas podem apresentar características intermediárias ou combinar regiões globulares e fibrosas em uma mesma molécula. Essa modularidade estrutural amplia ainda mais a capacidade funcional das proteínas, permitindo a integração de propriedades distintas em sistemas biológicos complexos.
Portanto, a distinção entre proteínas globulares e fibrosas não deve ser vista apenas como uma categorização morfológica, mas como uma expressão direta da relação entre estrutura e função. Essa relação é um dos princípios centrais da bioquímica e se manifesta de forma clara nesse contraste: enquanto a compactação estrutural favorece funções dinâmicas e específicas, a organização repetitiva e alongada sustenta funções mecânicas e estruturais.
6.11 Relação estrutura–função em proteínas #
A relação entre estrutura e função constitui um dos princípios mais fundamentais da bioquímica. Em proteínas, essa relação não é apenas conceitual, mas determinística: a função de uma proteína é diretamente condicionada por sua conformação tridimensional, que, por sua vez, deriva da sequência de aminoácidos. Essa hierarquia — sequência → estrutura → função — define a lógica molecular que governa praticamente todos os processos biológicos.
No nível mais básico, a função proteica depende da capacidade de interagir de forma específica com outras moléculas. Essa especificidade emerge da geometria e das propriedades químicas de regiões particulares da proteína, frequentemente organizadas em sítios de ligação ou sítios ativos. Nesses locais, a disposição espacial dos aminoácidos permite a formação de interações altamente seletivas, como ligações de hidrogênio, interações hidrofóbicas e forças eletrostáticas. Pequenas alterações na conformação desses sítios podem comprometer drasticamente a função, evidenciando o grau de precisão estrutural necessário para a atividade biológica.
Um exemplo emblemático dessa relação é observado em enzimas, nas quais o sítio ativo é moldado de forma a reconhecer o substrato com alta especificidade. Esse reconhecimento não é rigidamente estático, mas frequentemente envolve ajustes conformacionais, descritos pelo modelo de “ajuste induzido”. Nesse modelo, a ligação do substrato promove mudanças estruturais na enzima, otimizando a interação e favorecendo a catálise. Esse comportamento reforça a ideia de que a função proteica depende não apenas da estrutura estática, mas também de sua capacidade de adaptação dinâmica.
Além da catálise, a relação estrutura–função é evidente em proteínas de transporte, nas quais canais ou cavidades internas permitem a passagem seletiva de moléculas ou íons. Nesses casos, a arquitetura estrutural define não apenas o que pode ser transportado, mas também a direção e a eficiência do processo. De forma semelhante, proteínas de reconhecimento molecular, como anticorpos, apresentam regiões altamente variáveis cuja estrutura tridimensional determina a capacidade de reconhecer antígenos específicos.
A função proteica também pode depender da organização em níveis superiores, como a estrutura quaternária. Em proteínas oligoméricas, a interação entre subunidades permite fenômenos como cooperatividade e alosteria, nos quais mudanças conformacionais em uma parte da proteína influenciam outras regiões. Esse tipo de regulação é essencial para o controle de processos metabólicos, permitindo respostas rápidas e integradas às condições celulares.
Outro aspecto crítico é que a função proteica não depende apenas da estrutura global, mas também da presença de resíduos específicos em posições estratégicas. Aminoácidos com propriedades químicas particulares podem atuar como doadores ou aceitadores de prótons, nucleófilos ou estabilizadores de intermediários de reação. A posição tridimensional desses resíduos é determinante para sua participação em mecanismos catalíticos ou interações moleculares. Alterações pontuais na sequência, mesmo que não afetem drasticamente a estrutura global, podem comprometer essas funções locais.
A relação entre estrutura e função também se manifesta na sensibilidade das proteínas a alterações ambientais. Mudanças de pH, temperatura ou composição do meio podem afetar a estabilidade estrutural e, consequentemente, a atividade funcional. Em alguns casos, essas mudanças são exploradas biologicamente como mecanismos regulatórios; em outros, resultam em perda de função ou em patologias associadas à desnaturação ou ao mau dobramento proteico.
Do ponto de vista evolutivo, a conservação de estruturas tridimensionais é frequentemente mais significativa do que a conservação de sequências. Proteínas com sequências diferentes podem adotar estruturas semelhantes e desempenhar funções equivalentes, indicando que a forma tridimensional é o principal determinante funcional. Esse princípio é amplamente utilizado em bioinformática estrutural para prever funções de proteínas com base em sua estrutura.
Além disso, avanços recentes em biologia estrutural e modelagem computacional têm reforçado a centralidade dessa relação. A capacidade de prever estruturas tridimensionais a partir da sequência tem permitido inferir funções, identificar sítios de ligação e orientar o desenvolvimento de fármacos. Esses avanços evidenciam que compreender a estrutura de uma proteína é, em grande medida, compreender sua função.
Portanto, a relação estrutura–função em proteínas não é apenas um princípio descritivo, mas um eixo organizador da bioquímica. Ela estabelece que a atividade biológica emerge da organização espacial da matéria em nível molecular. Qualquer tentativa de compreender processos bioquímicos — seja catálise, transporte, sinalização ou regulação — exige, necessariamente, a análise integrada dessas duas dimensões. Ignorar essa relação compromete a interpretação dos mecanismos moleculares e limita a capacidade de intervenção em sistemas biológicos.
6.12 Introdução às proteínas funcionais #
A partir da consolidação dos níveis estruturais das proteínas, torna-se necessário avançar para uma questão central: como essas estruturas se traduzem em funções específicas dentro dos sistemas biológicos. As proteínas não são apenas entidades estruturais; são agentes ativos que executam, regulam e integram praticamente todos os processos celulares. A diversidade funcional observada decorre diretamente da diversidade estrutural previamente discutida, mas também da capacidade dessas macromoléculas de interagir de forma seletiva e dinâmica com outras moléculas.
As proteínas funcionais podem ser agrupadas em grandes classes, definidas de acordo com o tipo de atividade que desempenham. Entre as mais relevantes estão as proteínas catalíticas, conhecidas como enzimas. Essas moléculas aceleram reações químicas específicas, reduzindo a energia de ativação e permitindo que processos metabólicos ocorram em escalas de tempo compatíveis com a vida. A eficiência catalítica das enzimas depende da organização precisa de seus sítios ativos, onde resíduos de aminoácidos são posicionados de maneira a estabilizar estados de transição e facilitar transformações químicas.
Outra classe importante é composta pelas proteínas de transporte, responsáveis pela movimentação de moléculas e íons através de membranas ou no interior do organismo. Essas proteínas apresentam estruturas adaptadas à formação de canais, poros ou sítios de ligação que reconhecem seletivamente seus substratos. Em muitos casos, o transporte envolve mudanças conformacionais na proteína, permitindo a translocação de substâncias contra gradientes de concentração ou potencial eletroquímico.
As proteínas estruturais constituem um terceiro grupo fundamental. Diferentemente das enzimas e proteínas de transporte, sua função está relacionada à manutenção da integridade física das células e tecidos. Essas proteínas, frequentemente fibrosas, formam redes e matrizes que conferem resistência, elasticidade e organização espacial aos sistemas biológicos. Sua estabilidade estrutural é essencial para suportar forças mecânicas e manter a arquitetura celular.
Além dessas, destacam-se as proteínas regulatórias, que controlam a atividade de outras moléculas e processos celulares. Esse controle pode ocorrer por meio de interação direta com outras proteínas, modulação de vias metabólicas ou participação em cascatas de sinalização. Muitas proteínas regulatórias atuam como sensores moleculares, respondendo a mudanças no ambiente celular e ajustando a atividade metabólica de acordo com as necessidades do organismo.
As proteínas de defesa e reconhecimento molecular também desempenham papéis essenciais. Anticorpos, por exemplo, são capazes de reconhecer e neutralizar agentes estranhos com alta especificidade, enquanto outras proteínas participam de sistemas de identificação celular e resposta imune. Essa capacidade de reconhecimento depende de regiões estruturais altamente especializadas, que permitem a interação seletiva com alvos específicos.
Um aspecto crítico das proteínas funcionais é sua natureza dinâmica. Muitas dessas moléculas não operam como estruturas rígidas, mas como sistemas que alternam entre diferentes estados conformacionais. Essas transições são frequentemente induzidas pela ligação de ligantes, modificações químicas ou interações com outras proteínas. Essa plasticidade estrutural é essencial para funções como catálise, transporte e regulação, permitindo respostas rápidas e adaptativas às condições celulares.
Outro ponto relevante é que a função proteica raramente ocorre de forma isolada. Em sistemas biológicos, proteínas frequentemente atuam em conjunto, formando complexos multiproteicos ou participando de redes de interação. Essas associações ampliam a capacidade funcional, permitindo a integração de múltiplas atividades em processos coordenados. A organização em redes também facilita a regulação e a adaptação a diferentes contextos fisiológicos.
Do ponto de vista aplicado, a compreensão das proteínas funcionais é fundamental para áreas como biotecnologia, medicina e agronomia. Enzimas são amplamente utilizadas em processos industriais, proteínas de transporte são alvo de estudos farmacológicos, e proteínas regulatórias são centrais no desenvolvimento de terapias direcionadas. Em sistemas agrícolas, proteínas vegetais desempenham papéis críticos na resposta a estresses ambientais e na produtividade.
Portanto, a introdução às proteínas funcionais marca a transição entre a análise estrutural e a compreensão da atividade biológica. Ela evidencia que a estrutura, embora essencial, é apenas o meio pelo qual a função se manifesta. A partir desse ponto, a bioquímica passa a focar não apenas na forma das proteínas, mas naquilo que elas efetivamente fazem — e, principalmente, em como fazem — dentro da complexidade dos sistemas vivos.
6.13 Proteínas como sistemas dinâmicos #
A compreensão moderna das proteínas exige abandonar a visão clássica de estruturas rígidas e estáticas. Na realidade, proteínas são sistemas dinâmicos, capazes de explorar múltiplas conformações ao longo do tempo. Essa dinâmica estrutural não é um detalhe secundário, mas um componente essencial de sua função. Em muitos casos, a atividade biológica depende diretamente da capacidade da proteína de alternar entre diferentes estados conformacionais em resposta a estímulos específicos.
Do ponto de vista físico-químico, uma proteína não ocupa uma única conformação, mas um conjunto de estados estruturais energeticamente próximos, frequentemente descrito como um “ensemble conformacional”. Dentro desse conjunto, diferentes conformações coexistem em equilíbrio dinâmico, com probabilidades determinadas pela energia livre de cada estado. Pequenas variações no ambiente — como mudanças de pH, temperatura ou presença de ligantes — podem deslocar esse equilíbrio, favorecendo estados específicos que correspondem a diferentes funções [Figura].
Esse comportamento é particularmente evidente em enzimas, nas quais a dinâmica estrutural está diretamente relacionada à catálise. O modelo de ajuste induzido descreve como a ligação do substrato promove alterações conformacionais na proteína, otimizando a geometria do sítio ativo para facilitar a reação. No entanto, modelos mais recentes sugerem que essas conformações já existem previamente no ensemble da proteína, e que o ligante seleciona a conformação mais adequada — um conceito conhecido como “seleção conformacional”. Na prática, ambos os mecanismos podem coexistir, refletindo a complexidade do sistema.
A dinâmica proteica também é fundamental em processos de regulação alostérica. Em proteínas regulatórias, a ligação de uma molécula em um sítio específico pode induzir mudanças estruturais que se propagam ao longo da molécula, alterando a atividade em regiões distantes. Esse tipo de comunicação intramolecular depende da flexibilidade estrutural e da capacidade de reorganização das interações internas. Assim, a proteína atua como um sistema integrado, no qual diferentes regiões estão funcionalmente conectadas.
Outro exemplo relevante é observado em proteínas de transporte, que frequentemente operam por meio de ciclos conformacionais. Essas proteínas alternam entre estados estruturais distintos para permitir a passagem de moléculas através de membranas, muitas vezes contra gradientes de concentração. Cada estado conformacional apresenta afinidade diferente pelo substrato, garantindo a direcionalidade do processo. Sem essa capacidade de transição estrutural, o transporte seletivo seria inviável.
A dinâmica também se manifesta em regiões intrinsecamente desordenadas de proteínas. Diferentemente das regiões estruturadas, essas porções não adotam uma conformação fixa, permanecendo flexíveis e adaptáveis. Essa característica permite interações com múltiplos parceiros e participação em processos regulatórios complexos. Proteínas com regiões desordenadas são comuns em sistemas de sinalização e controle, onde a versatilidade funcional é mais importante do que a estabilidade estrutural.
Em termos moleculares, a dinâmica proteica ocorre em diferentes escalas de tempo e espaço. Movimentos rápidos, como vibrações de ligações químicas, coexistem com rearranjos mais lentos, como mudanças conformacionais globais. Essa hierarquia de movimentos permite que a proteína responda de maneira eficiente a diferentes estímulos, ajustando sua estrutura conforme necessário para desempenhar sua função.
Do ponto de vista experimental, a investigação da dinâmica proteica requer técnicas capazes de capturar não apenas estruturas estáticas, mas também movimentos ao longo do tempo. Métodos como ressonância magnética nuclear (RMN), espectroscopia de fluorescência e simulações de dinâmica molecular têm sido fundamentais para revelar esses comportamentos. Esses estudos mostram que a função proteica está frequentemente associada a transições entre estados, e não a uma única conformação estável.
A implicação prática dessa visão é significativa. Em áreas como o desenvolvimento de fármacos, por exemplo, considerar apenas a estrutura estática de uma proteína pode ser insuficiente. A identificação de estados conformacionais alternativos pode revelar novos sítios de ligação e oportunidades terapêuticas. Da mesma forma, na engenharia de proteínas, a modulação da flexibilidade estrutural pode ser utilizada para otimizar atividade, estabilidade ou especificidade.
Portanto, as proteínas devem ser entendidas como sistemas dinâmicos, nos quais estrutura e movimento estão intrinsecamente ligados. A função não emerge apenas da forma, mas da capacidade de transitar entre formas. Essa perspectiva amplia a compreensão da bioquímica, deslocando o foco de estruturas fixas para sistemas em constante adaptação, capazes de responder de maneira precisa e eficiente às demandas do ambiente biológico.
6.14 Erros de dobramento e doenças #
A fidelidade do dobramento proteico é um requisito crítico para a manutenção da homeostase celular. Embora a sequência de aminoácidos contenha a informação necessária para a aquisição da conformação nativa, o processo de enovelamento está sujeito a falhas. Quando isso ocorre, a proteína pode assumir estruturas incorretas, parcialmente dobradas ou agregadas, comprometendo sua função e, em muitos casos, desencadeando efeitos patológicos. Assim, os erros de dobramento não são eventos marginais, mas componentes centrais na etiologia de diversas doenças.
Do ponto de vista molecular, proteínas mal dobradas apresentam frequentemente regiões hidrofóbicas expostas que, em condições normais, estariam enterradas no interior da estrutura. Essa exposição favorece interações não específicas entre diferentes moléculas, levando à formação de agregados. Esses agregados podem assumir formas amorfas ou altamente organizadas, como as fibrilas amiloides, caracterizadas por estruturas ricas em folhas β empilhadas [Figura]. A formação dessas estruturas está associada a diversas doenças neurodegenerativas, nas quais o acúmulo progressivo de agregados compromete a função celular.
Entre os exemplos mais estudados estão doenças como Alzheimer, Parkinson e Huntington, todas marcadas pela deposição de proteínas mal conformadas em tecidos específicos. No caso do Alzheimer, a agregação do peptídeo β-amiloide e da proteína tau leva à formação de placas e emaranhados neurofibrilares, interferindo na comunicação neuronal. Já na doença de Parkinson, a proteína α-sinucleína forma agregados que afetam a função dos neurônios dopaminérgicos. Esses processos evidenciam que a toxicidade não decorre apenas da perda de função da proteína original, mas também do ganho de função tóxica associado aos agregados.
Um caso particular de erro de dobramento é observado nas doenças priônicas. Nesses sistemas, uma proteína normal pode ser convertida em uma forma patológica por meio de interação direta com uma proteína já mal dobrada. Essa conversão envolve uma mudança conformacional que favorece a formação de estruturas ricas em folhas β, altamente estáveis e resistentes à degradação. O aspecto singular desse mecanismo é sua natureza autocatalítica, na qual a forma patológica induz a conversão de outras moléculas, levando à propagação do estado anômalo sem necessidade de material genético.
As células possuem mecanismos sofisticados para lidar com proteínas mal dobradas. Sistemas de controle de qualidade, localizados principalmente no retículo endoplasmático e no citosol, monitoram o estado conformacional das proteínas recém-sintetizadas. Proteínas que não atingem a conformação adequada são reconhecidas por chaperonas moleculares, que tentam promover seu dobramento correto. Quando essa tentativa falha, as proteínas são direcionadas para sistemas de degradação, como o proteassoma ou a autofagia. Esse conjunto de processos constitui a chamada proteostase, essencial para a manutenção do equilíbrio proteico celular.
Entretanto, quando a capacidade desses sistemas é excedida — seja por mutações que aumentam a propensão ao mau dobramento, seja por condições ambientais adversas — ocorre o acúmulo de proteínas defeituosas. Esse acúmulo pode desencadear respostas celulares de estresse, como a resposta a proteínas mal dobradas (UPR), que tenta restaurar o equilíbrio, mas que, em situações prolongadas, pode levar à morte celular.
Além das doenças neurodegenerativas, erros de dobramento estão envolvidos em diversas outras patologias. Em alguns casos, mutações levam à produção de proteínas que não conseguem atingir a conformação funcional e são degradadas prematuramente, resultando em deficiência funcional. Em outros, proteínas parcialmente dobradas podem se acumular em compartimentos celulares, interferindo em processos fisiológicos normais.
Do ponto de vista aplicado, a compreensão dos mecanismos de dobramento incorreto abre possibilidades terapêuticas. Estratégias incluem o desenvolvimento de moléculas capazes de estabilizar a conformação nativa, impedir a agregação ou promover a degradação seletiva de proteínas mal dobradas. Essas abordagens têm sido exploradas no tratamento de doenças associadas à agregação proteica, embora ainda enfrentem desafios significativos.
Portanto, os erros de dobramento representam uma interface crítica entre bioquímica estrutural e patologia. Eles evidenciam que a função proteica depende não apenas da sequência e da estrutura, mas também da capacidade do sistema celular de garantir o dobramento correto e de eliminar formas defeituosas. Quando esse equilíbrio é perdido, as consequências se manifestam em nível molecular, celular e sistêmico, reforçando a importância do controle estrutural das proteínas para a saúde dos organismos.
6.15 Aplicações em bioquímica aplicada #
A consolidação do conhecimento sobre aminoácidos, estrutura e dinâmica das proteínas não tem apenas valor conceitual; ela sustenta um amplo conjunto de aplicações práticas em áreas estratégicas como saúde, agricultura, indústria e biotecnologia. Nesse contexto, as proteínas deixam de ser apenas objetos de estudo e passam a ser ferramentas tecnológicas, cuja manipulação racional permite resolver problemas concretos e gerar valor econômico.
No campo da biotecnologia, as proteínas — especialmente as enzimas — são utilizadas como catalisadores em processos industriais. Diferentemente de catalisadores químicos convencionais, enzimas operam sob condições mais brandas de temperatura e pH, apresentam alta especificidade e geram menos subprodutos indesejados. Esses atributos são explorados em setores como indústria alimentícia, farmacêutica e de biocombustíveis. A compreensão da relação estrutura–função permite otimizar essas enzimas por meio de engenharia de proteínas, ajustando sua estabilidade, atividade e especificidade para condições industriais específicas.
A engenharia de proteínas, em particular, representa um avanço direto do entendimento estrutural desenvolvido ao longo deste capítulo. A partir da modificação da sequência de aminoácidos, é possível alterar propriedades funcionais de maneira direcionada. Técnicas como mutagênese sítio-dirigida permitem substituir resíduos específicos para melhorar a estabilidade térmica, aumentar a eficiência catalítica ou modificar a afinidade por substratos. Em abordagens mais recentes, a evolução dirigida combina mutações aleatórias com seleção funcional, simulando processos evolutivos em escala acelerada para gerar proteínas com características desejadas.
Na área da saúde, proteínas são centrais tanto como alvos terapêuticos quanto como agentes terapêuticos. Fármacos modernos frequentemente são projetados para interagir com proteínas específicas, como enzimas ou receptores, modulando sua atividade. A compreensão detalhada da estrutura tridimensional dessas proteínas permite identificar sítios de ligação e desenvolver moléculas com alta afinidade e especificidade. Paralelamente, proteínas terapêuticas — como anticorpos monoclonais, hormônios recombinantes e enzimas substitutivas — são utilizadas diretamente no tratamento de diversas doenças. Nesses casos, a estabilidade estrutural e a correta conformação são determinantes para a eficácia clínica.
No contexto agrícola, as proteínas desempenham papel estratégico tanto na produção quanto na proteção de culturas. Enzimas vegetais estão diretamente envolvidas em processos como crescimento, metabolismo e resposta a estresses ambientais. A manipulação genética de plantas para expressar proteínas específicas pode aumentar resistência a pragas, tolerância à seca e eficiência no uso de nutrientes. Além disso, proteínas de origem microbiana, como toxinas específicas, são utilizadas no controle biológico de pragas, reduzindo a dependência de pesticidas químicos.
A bioquímica aplicada também se estende à produção de alimentos. Proteínas são responsáveis por propriedades sensoriais e funcionais, como textura, solubilidade e capacidade de emulsificação. A modificação dessas propriedades, por meio de processos enzimáticos ou engenharia molecular, permite desenvolver alimentos com características específicas, como maior valor nutricional ou melhor estabilidade. No seu contexto específico, o desenvolvimento de proteínas vegetais — como concentrados proteicos — depende diretamente da compreensão das propriedades estruturais e químicas dessas macromoléculas.
Outro campo relevante é o diagnóstico molecular, no qual proteínas são utilizadas como biomarcadores ou componentes de sistemas analíticos. Enzimas, anticorpos e proteínas de ligação específica são empregados em testes laboratoriais para detectar a presença de substâncias em amostras biológicas. A precisão desses testes depende da especificidade e estabilidade das proteínas envolvidas, reforçando a importância do controle estrutural.
Do ponto de vista industrial e tecnológico, a estabilidade proteica é um fator crítico. Processos que envolvem variações de temperatura, pH ou presença de solventes exigem proteínas capazes de manter sua função sob condições adversas. Isso impulsiona o desenvolvimento de proteínas mais robustas, seja por seleção natural de organismos extremófilos, seja por engenharia racional. A capacidade de projetar proteínas com estabilidade aumentada é um diferencial competitivo em diversos setores.
Entretanto, é necessário reconhecer um ponto estratégico: conhecimento sem foco aplicado tende a se dispersar. O domínio estrutural das proteínas só se traduz em vantagem prática quando direcionado a problemas específicos. No seu contexto — agro, biotecnologia e inovação — o ponto de maior alavancagem está na interseção entre proteínas funcionais + aplicação prática + escalabilidade. Qualquer esforço fora desse eixo tende a gerar baixo retorno.
Portanto, as aplicações em bioquímica aplicada representam a materialização do conhecimento estrutural em soluções reais. Elas evidenciam que compreender proteínas não é apenas um exercício acadêmico, mas um caminho direto para inovação tecnológica, geração de valor e impacto econômico. A partir desse ponto, o desafio deixa de ser entender as proteínas e passa a ser utilizá-las estrategicamente.
Perguntas de estudo dirigido — Capítulo 6: Aminoácidos, Estrutura de Proteínas e Relação Estrutura–Função #
- Explique a estrutura geral de um aminoácido.
Identifique o carbono α, o grupo amino, o grupo carboxila, o hidrogênio e a cadeia lateral R. Em seguida, explique por que a cadeia lateral é o principal fator responsável pelas diferenças químicas entre os aminoácidos. - Como o pH influencia o estado de ionização dos aminoácidos?
Descreva a forma catiônica, zwitteriônica e aniônica de um aminoácido simples. Relacione esses estados aos conceitos de pKa e ponto isoelétrico. - Descreva a formação da ligação peptídica.
Explique a reação de condensação entre dois aminoácidos, a liberação de água, o caráter parcial de dupla ligação da ligação peptídica e a importância da direção N-terminal → C-terminal. - Compare os quatro níveis de organização estrutural das proteínas.
Diferencie estrutura primária, secundária, terciária e quaternária, indicando quais interações químicas predominam em cada nível. - Por que a estrutura tridimensional de uma proteína é essencial para sua função?
Use exemplos como enzimas, hemoglobina, proteínas fibrosas ou proteínas desnaturadas para explicar a relação entre sequência, dobramento, estabilidade e atividade biológica.
Pergunta para estimular pesquisa #
- Pesquise uma doença associada ao mau dobramento ou agregação de proteínas — como Alzheimer, Parkinson, Huntington, anemia falciforme ou doenças priônicas — e explique qual proteína está envolvida, qual alteração estrutural ocorre e como essa alteração compromete a função celular ou fisiológica.
Referências #
NELSON, David L.; COX, Michael M. Princípios de bioquímica de Lehninger. 6. ed. Porto Alegre: Artmed, 2014.
STRYER, Lubert; BERG, Jeremy M.; TYMOCZKO, John L.; GATTO JUNIOR, Gregory J. Bioquímica. 8. ed. Rio de Janeiro: Guanabara Koogan, 2014.
VOET, Donald; VOET, Judith G.; PRATT, Charlotte W. Fundamentos de bioquímica: a vida em nível molecular. 4. ed. Porto Alegre: Artmed, 2014.
MURRAY, Robert K. et al. Bioquímica ilustrada de Harper. 30. ed. Porto Alegre: AMGH, 2017.



