8.1 — O citoesqueleto como sistema estrutural dinâmico da célula #
A organização interna das células eucariontes revela um paradoxo funcional que desafia interpretações simplistas da estrutura biológica. A célula precisa, simultaneamente, manter sua forma, resistir a tensões mecânicas, organizar seus componentes internos e, ao mesmo tempo, ser altamente dinâmica, capaz de se deformar, migrar, dividir-se e responder rapidamente a estímulos ambientais. Essa dualidade entre estabilidade e plasticidade não pode ser explicada apenas pela presença de membranas ou pela composição química do citoplasma. Ela emerge de um sistema estrutural altamente organizado e regulado: o citoesqueleto.
Longe de ser uma “armação passiva”, como sugeriam interpretações iniciais baseadas em limitações técnicas da microscopia óptica, o citoesqueleto constitui uma rede tridimensional de filamentos proteicos que permeia todo o citoplasma e estabelece uma arquitetura funcional integrada. Essa rede não apenas sustenta a forma celular, mas também define a posição de organelas, orienta o tráfego intracelular e participa ativamente de processos fundamentais como divisão celular, endocitose, secreção e motilidade. [Figura]
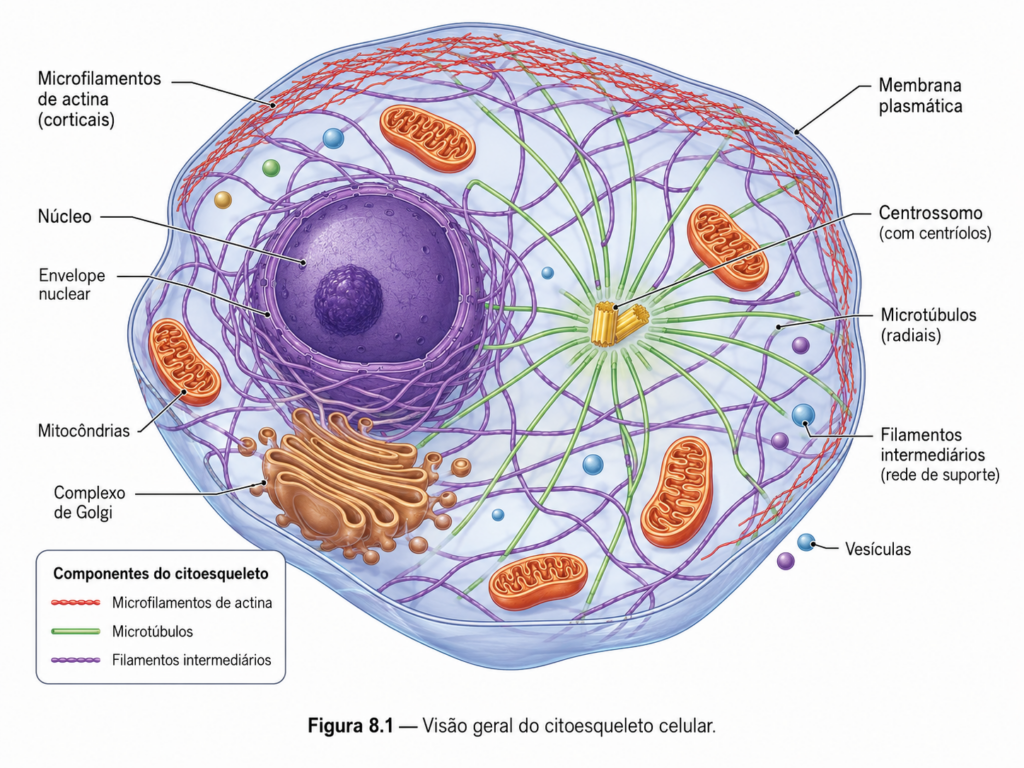
Do ponto de vista molecular, o citoesqueleto é composto por diferentes classes de filamentos proteicos que apresentam propriedades físico-químicas distintas, permitindo a execução de funções específicas e complementares. Essa diversidade estrutural não é redundante; ao contrário, ela é essencial para que a célula opere como um sistema adaptativo. A coexistência de elementos mais rígidos, capazes de resistir a deformações, com estruturas altamente dinâmicas, sujeitas a rápida montagem e desmontagem, estabelece a base para o comportamento mecânico da célula em diferentes contextos fisiológicos.
Um aspecto central do citoesqueleto é sua natureza dinâmica. Diferentemente de estruturas estáticas, os filamentos citosqueléticos estão em constante reorganização, por meio de processos regulados de polimerização e despolimerização. Essa dinâmica permite que a célula altere rapidamente sua forma e reorganize sua estrutura interna em resposta a sinais extracelulares ou demandas metabólicas. Em células em migração, por exemplo, regiões específicas do citoesqueleto são continuamente remodeladas para promover protrusões da membrana plasmática, enquanto outras regiões se contraem para impulsionar o deslocamento celular. [Figura]
Essa propriedade dinâmica é fortemente dependente de interações com proteínas acessórias que regulam a nucleação, o crescimento, a estabilização e a fragmentação dos filamentos. Essas proteínas funcionam como elementos de controle que ajustam o comportamento do citoesqueleto de acordo com o estado fisiológico da célula. Dessa forma, o citoesqueleto não atua isoladamente, mas como parte de uma rede regulatória complexa que integra sinais bioquímicos e mecânicos.
A capacidade do citoesqueleto de gerar força mecânica representa outro aspecto fundamental de sua função. Por meio da interação com proteínas motoras, que convertem energia química derivada da hidrólise de ATP em trabalho mecânico, o citoesqueleto participa diretamente do transporte intracelular de vesículas, organelas e complexos macromoleculares. Esse transporte não ocorre por difusão aleatória, mas sim de forma direcionada e eficiente ao longo de trilhos definidos por filamentos citosqueléticos. [Figura]
Além do transporte intracelular, a geração de força pelo citoesqueleto é essencial para processos como contração celular, formação do fuso mitótico e segregação cromossômica durante a divisão celular. Nesse contexto, o citoesqueleto atua como um sistema ativo capaz de converter energia em movimento e organização espacial, estabelecendo uma relação direta entre estrutura molecular e função biológica.
A integração do citoesqueleto com a membrana plasmática e com outros sistemas celulares amplia ainda mais sua relevância funcional. Elementos do citoesqueleto interagem com proteínas de membrana e complexos de adesão, permitindo que forças mecânicas sejam transmitidas entre o interior da célula e o ambiente extracelular. Essa interação é essencial para processos como adesão celular, percepção de estímulos mecânicos e resposta a variações no microambiente, incluindo alterações de pressão, rigidez do substrato e disponibilidade de nutrientes.
Em sistemas vegetais, essa integração assume papel estratégico na resposta a estresses ambientais. Alterações na organização do citoesqueleto estão associadas a respostas adaptativas a condições como déficit hídrico, salinidade e variações térmicas, influenciando diretamente processos de crescimento, expansão celular e orientação do desenvolvimento. [Quadro]
Do ponto de vista conceitual, o citoesqueleto deve ser compreendido não apenas como um conjunto de estruturas filamentosas, mas como um sistema funcional integrado que conecta forma, movimento, organização interna e resposta ao ambiente. Essa perspectiva rompe com abordagens fragmentadas da célula e reforça a ideia de que os componentes celulares operam em rede, com interdependência estrutural e funcional.
A compreensão do citoesqueleto como um sistema dinâmico e regulado estabelece a base para a análise detalhada de seus componentes, que serão explorados nas seções seguintes. A diversidade de filamentos, suas propriedades específicas e seus mecanismos de regulação revelam como a célula constrói, mantém e remodela sua própria arquitetura, garantindo eficiência funcional em um ambiente em constante mudança.
8.2 — Organização molecular do citoesqueleto #
A compreensão do citoesqueleto como um sistema funcional exige avançar da visão global para o nível molecular, onde se estabelece a base físico-química de sua organização e dinâmica. A aparente complexidade estrutural observada no interior celular resulta, na realidade, de um conjunto relativamente restrito de proteínas capazes de se auto-organizar em polímeros altamente regulados. Essa capacidade de auto-organização, aliada a mecanismos de controle finos e responsivos, confere ao citoesqueleto a propriedade essencial de ser simultaneamente estruturado e adaptativo.
No núcleo dessa organização estão proteínas globulares que possuem a capacidade intrínseca de polimerizar, formando filamentos alongados com propriedades mecânicas específicas. Esse processo não ocorre de maneira espontânea e desordenada no ambiente celular, mas segue princípios termodinâmicos e cinéticos rigorosamente modulados por condições locais, como concentração de monômeros, presença de cofatores e interação com proteínas reguladoras. A célula, portanto, controla o citoesqueleto não apenas pela síntese de seus componentes, mas principalmente pela regulação de sua montagem e desmontagem.
[Figura]
8.2.1 — Proteínas estruturais e polimerização #
Os principais componentes do citoesqueleto são formados por proteínas capazes de alternar entre estados monoméricos e poliméricos, estabelecendo um equilíbrio dinâmico que define a arquitetura celular. Essas proteínas apresentam domínios estruturais que favorecem interações específicas, permitindo a formação de polímeros lineares ou associações laterais mais complexas. A natureza dessas interações determina propriedades como rigidez, flexibilidade e resistência à tensão.
A polimerização ocorre por meio da adição sequencial de subunidades proteicas a uma extremidade do filamento, frequentemente acompanhada de mudanças conformacionais induzidas por nucleotídeos como ATP ou GTP. Esse acoplamento entre estado energético e organização estrutural permite que o citoesqueleto seja sensível ao estado metabólico da célula. Em condições de alta disponibilidade energética, por exemplo, a montagem de filamentos pode ser favorecida, enquanto situações de estresse energético tendem a promover sua desorganização.
A nucleação representa um ponto crítico nesse processo. A formação inicial de um núcleo estável de polimerização é energeticamente desfavorável, o que impede a formação indiscriminada de filamentos. A célula contorna essa limitação por meio de proteínas especializadas que atuam como plataformas de nucleação, garantindo que a formação de novos filamentos ocorra em locais específicos e de maneira controlada. Esse mecanismo assegura que a organização do citoesqueleto seja espacialmente definida, evitando interferências com outras estruturas celulares.
[Figura]
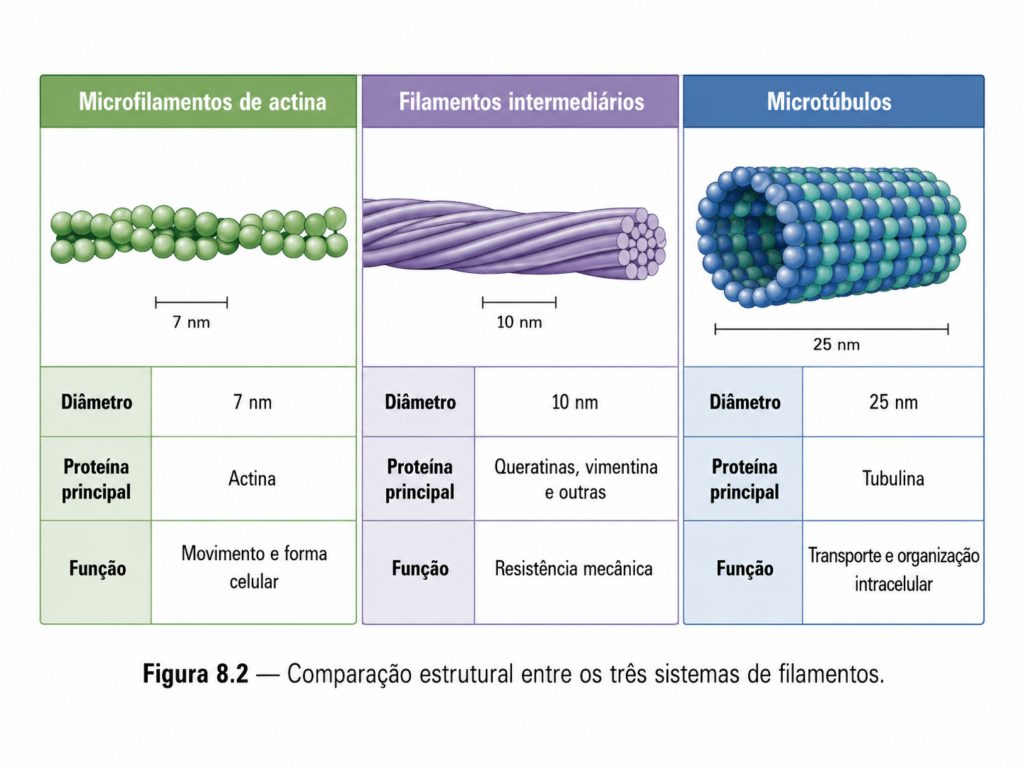
8.2.2 — Dinâmica de montagem e desmontagem #
A característica mais marcante do citoesqueleto reside em sua dinâmica contínua, sustentada por ciclos de montagem e desmontagem que ocorrem em escalas de tempo compatíveis com as necessidades funcionais da célula. Diferentemente de estruturas estáticas, os filamentos citosqueléticos estão em constante renovação, permitindo ajustes rápidos na organização celular.
Esse comportamento dinâmico é governado por fenômenos como o crescimento diferencial das extremidades dos filamentos, frequentemente descrito em termos de polaridade estrutural. Uma extremidade tende a apresentar maior taxa de adição de subunidades, enquanto a outra pode favorecer a dissociação. Esse desequilíbrio cria um fluxo contínuo de subunidades ao longo do filamento, mesmo na ausência de mudanças no comprimento total, fenômeno conhecido como “treadmilling”. [Figura]
Em sistemas baseados em tubulina, observa-se um comportamento adicional denominado instabilidade dinâmica, caracterizado por fases alternadas de crescimento e rápida despolimerização. Esse mecanismo permite a exploração rápida do espaço intracelular, sendo particularmente relevante durante a formação do fuso mitótico, onde microtúbulos precisam localizar e capturar cromossomos com alta eficiência.
A desmontagem dos filamentos não representa um processo meramente passivo, mas é ativamente regulada por proteínas que promovem a fragmentação ou desestabilização das estruturas existentes. Esse controle garante que a reorganização do citoesqueleto ocorra de maneira coordenada, evitando colapsos estruturais e permitindo respostas rápidas a sinais internos e externos.
8.2.3 — Regulação espacial e temporal do citoesqueleto #
A organização do citoesqueleto não depende apenas das propriedades intrínsecas de seus componentes, mas de um sistema de regulação que integra sinais bioquímicos e mecânicos para definir onde e quando os filamentos devem se formar ou se desestruturar. Essa regulação é essencial para a manutenção da polaridade celular, para a execução de processos direcionados e para a adaptação a condições ambientais variáveis.
Proteínas reguladoras atuam como sensores e efetores, respondendo a sinais provenientes de receptores de membrana, vias de transdução de sinal e alterações no estado fisiológico da célula. Essas proteínas controlam processos como nucleação, alongamento, estabilização e ancoragem dos filamentos, estabelecendo padrões organizacionais específicos para cada tipo celular.
[Figura]
A distribuição espacial do citoesqueleto está intimamente associada à compartimentalização funcional da célula. Regiões envolvidas em processos de migração, por exemplo, apresentam intensa atividade de polimerização de actina na periferia celular, enquanto áreas mais internas tendem a concentrar estruturas associadas ao transporte intracelular. Essa organização diferencial permite que a célula execute múltiplas funções simultaneamente sem comprometer sua integridade estrutural.
Do ponto de vista temporal, a reorganização do citoesqueleto acompanha eventos celulares específicos, como progressão do ciclo celular, resposta a estímulos externos e processos de diferenciação. Durante a divisão celular, por exemplo, ocorre uma reconfiguração profunda da arquitetura citosquelética, com desmontagem de estruturas pré-existentes e formação de novas configurações especializadas, como o fuso mitótico e o anel contrátil.
Em contextos aplicados, especialmente em sistemas vegetais, a regulação do citoesqueleto está associada a processos de crescimento orientado, deposição de parede celular e resposta a estresses ambientais. Alterações na organização dos filamentos podem influenciar diretamente a direção do crescimento celular e a eficiência na absorção de recursos, estabelecendo conexões claras entre biologia celular e produtividade agrícola. [Quadro]
A análise da organização molecular do citoesqueleto evidencia que sua funcionalidade emerge da integração entre propriedades estruturais, dinâmica regulada e controle espacial preciso. Essa base conceitual é essencial para compreender como diferentes tipos de filamentos contribuem de maneira específica para a arquitetura e o comportamento celular, tema que será aprofundado nas seções seguintes.
8.3 — Microfilamentos de actina e controle da forma celular #
A definição da forma celular e sua capacidade de se modificar ao longo do tempo dependem de um conjunto de estruturas altamente organizadas que operam na interface entre estabilidade estrutural e plasticidade dinâmica. Entre essas estruturas, os microfilamentos de actina ocupam uma posição central, não apenas por sua abundância no citoplasma, mas pela versatilidade funcional que apresentam. A actina permite que a célula mantenha integridade mecânica ao mesmo tempo em que promove deformações controladas, essenciais para processos como migração, adesão e divisão celular.
Os microfilamentos formam uma rede densa e altamente dinâmica, especialmente concentrada na região cortical, imediatamente abaixo da membrana plasmática. Essa localização estratégica permite que a actina exerça controle direto sobre a forma celular, modulando tensões superficiais e organizando domínios funcionais específicos. Em células em repouso, essa rede contribui para a manutenção da arquitetura celular; em células ativas, torna-se um sistema altamente responsivo, capaz de reorganizar-se em questão de segundos. [Figura]
8.3.1 — Estrutura e polaridade dos filamentos de actina #
A actina é uma proteína globular que, ao polimerizar, forma filamentos finos e helicoidais com propriedades estruturais bem definidas. Esses filamentos apresentam polaridade intrínseca, caracterizada por duas extremidades estruturalmente distintas, frequentemente denominadas extremidade “positiva” e extremidade “negativa”. Essa polaridade não é apenas uma característica geométrica, mas um elemento funcional essencial que determina a direção do crescimento dos filamentos e a interação com proteínas motoras.
A polimerização da actina ocorre pela adição de monômeros em uma das extremidades, geralmente com maior eficiência na extremidade positiva. Esse crescimento assimétrico estabelece um fluxo direcional de subunidades ao longo do filamento, criando uma base estrutural para processos que exigem orientação espacial, como transporte intracelular e formação de protrusões celulares.
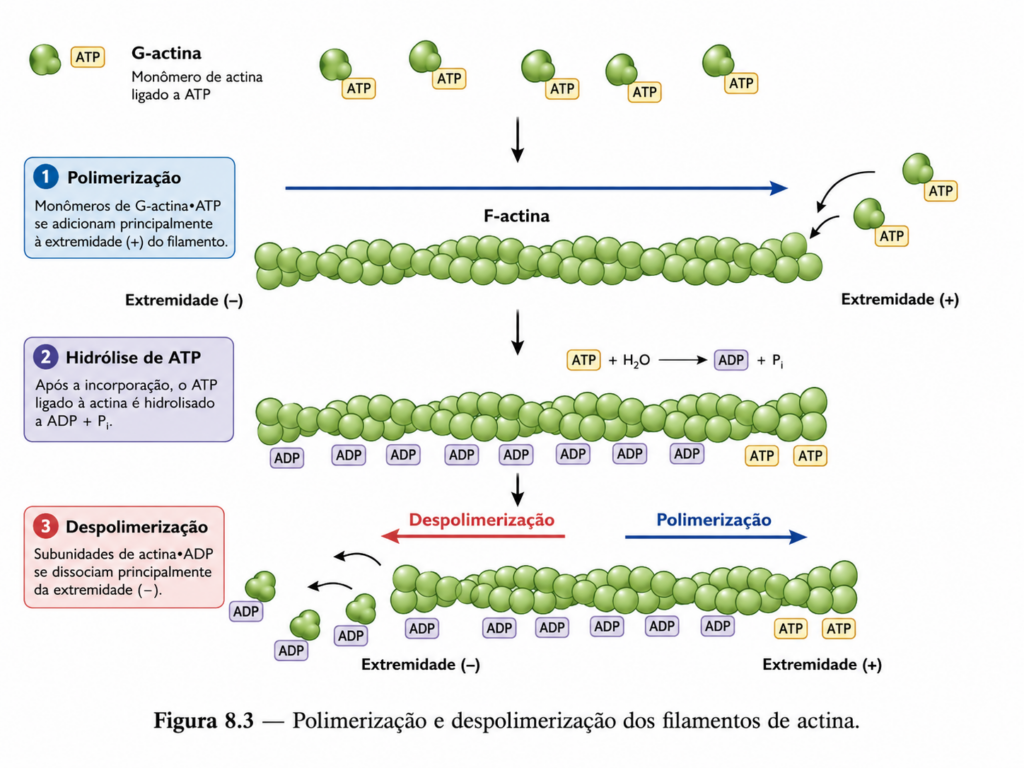
A estabilidade dos filamentos depende do estado de ligação a nucleotídeos. A actina associada a ATP tende a favorecer a polimerização, enquanto a hidrólise subsequente para ADP altera a conformação da subunidade, tornando o filamento mais suscetível à despolimerização. Esse ciclo bioquímico conecta diretamente o metabolismo energético à organização estrutural, permitindo que a célula ajuste a dinâmica do citoesqueleto de acordo com sua condição fisiológica.
8.3.2 — Proteínas associadas à actina #
A funcionalidade dos microfilamentos não pode ser compreendida apenas pela análise da actina isoladamente. Um amplo conjunto de proteínas associadas regula sua organização, definindo padrões específicos de montagem, estabilidade e interação com outros componentes celulares. Essas proteínas atuam em diferentes níveis, desde a nucleação inicial até a organização em estruturas complexas.
Proteínas de nucleação controlam o surgimento de novos filamentos, garantindo que a polimerização ocorra em regiões específicas da célula. Outras proteínas promovem a ligação lateral entre filamentos, formando feixes ou redes tridimensionais que conferem diferentes propriedades mecânicas ao citoesqueleto. Estruturas mais rígidas, como feixes paralelos, são frequentemente associadas a funções de sustentação, enquanto redes mais ramificadas estão relacionadas à geração de força e movimento.
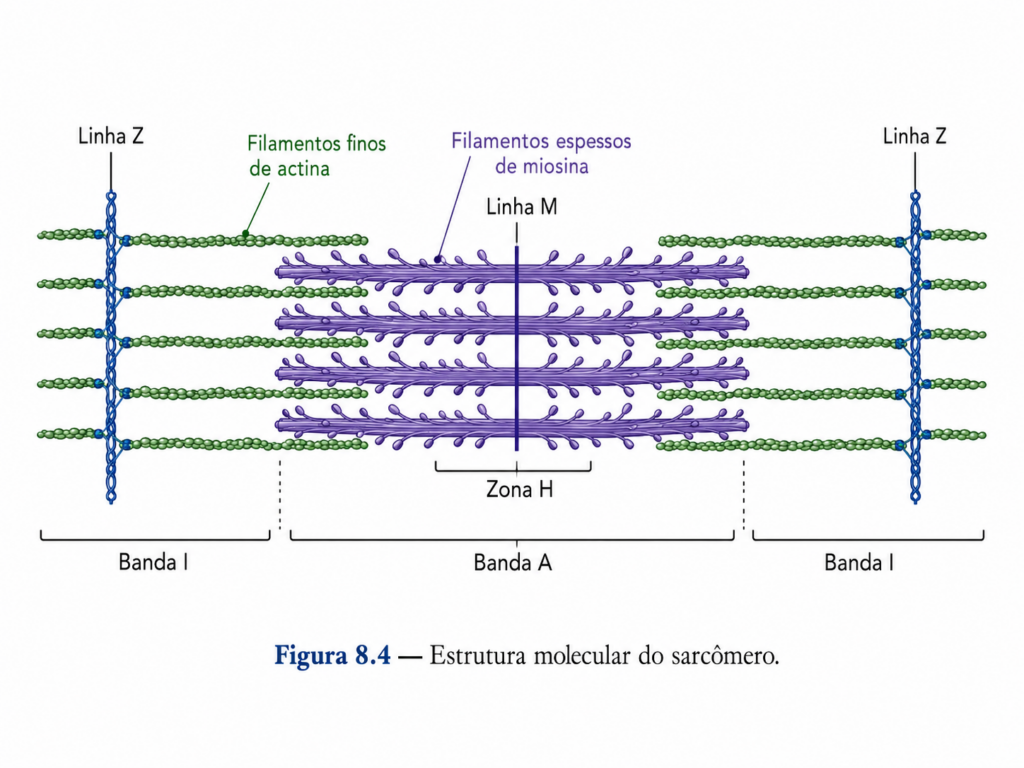
Além disso, proteínas reguladoras controlam o comprimento dos filamentos, promovendo tanto sua estabilização quanto sua fragmentação. Esse controle permite ajustes finos na arquitetura celular, evitando crescimento desordenado e garantindo renovação contínua da rede de actina. Proteínas de ligação à membrana também desempenham papel fundamental, ancorando os filamentos à superfície celular e permitindo a transmissão de forças mecânicas.
8.3.3 — Actina na contração e na morfologia celular #
Um dos aspectos mais relevantes da actina é sua capacidade de participar diretamente da geração de força mecânica. Essa função é mediada pela interação com proteínas motoras, especialmente as miosinas, que utilizam energia química para produzir movimento relativo entre filamentos. Esse mecanismo é amplamente conhecido em células musculares, mas também ocorre em diversos contextos celulares não especializados, onde contribui para processos como contração cortical, transporte intracelular e reorganização estrutural.
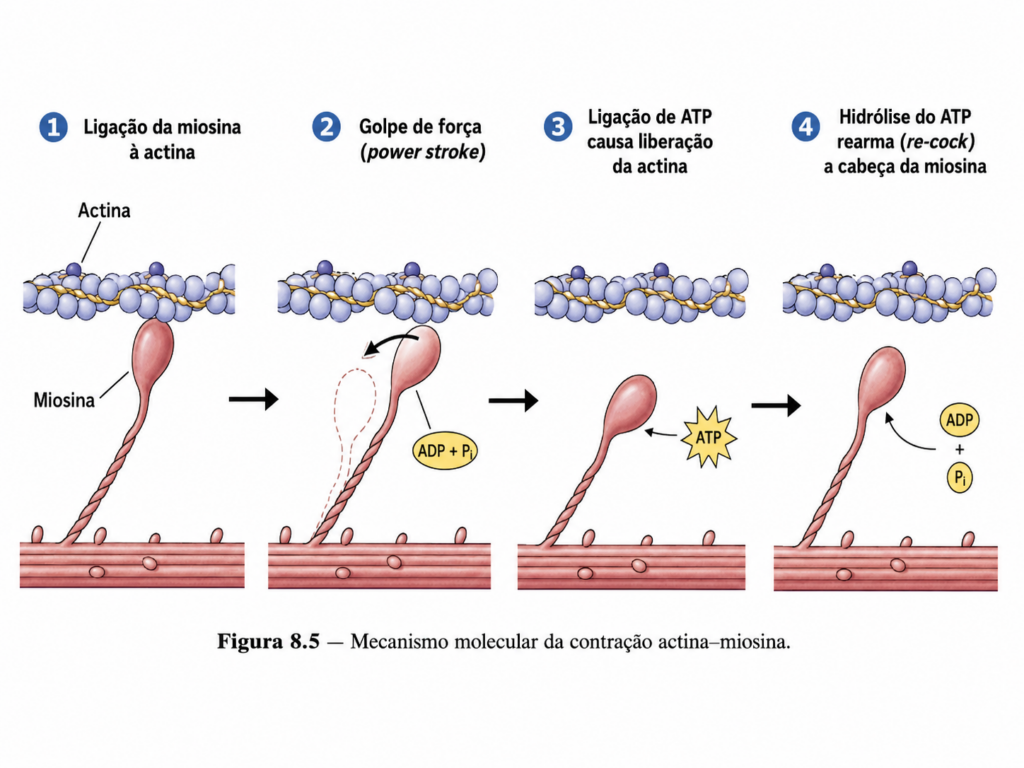
Na região cortical, a interação entre actina e miosina gera tensões que regulam a forma celular e a rigidez da membrana plasmática. Esse sistema contrátil permite que a célula ajuste sua superfície em resposta a estímulos mecânicos e químicos, controlando processos como endocitose, exocitose e formação de junções celulares.
A actina também desempenha papel central na formação de estruturas especializadas associadas à motilidade celular. Protrusões como lamelipódios e filopódios são geradas por polimerização direcionada de actina, que empurra a membrana plasmática para fora, permitindo que a célula explore seu ambiente e estabeleça novos pontos de adesão. Esse processo é altamente coordenado, envolvendo ciclos de extensão, adesão e retração que resultam em deslocamento celular efetivo.
Em sistemas vegetais, a actina está envolvida na orientação do crescimento celular e no transporte intracelular de vesículas, especialmente em células com crescimento polarizado, como tubos polínicos e células radiculares. Alterações na organização da actina podem impactar diretamente a direção do crescimento e a capacidade da planta de responder a estímulos ambientais, como gravidade, luz e disponibilidade hídrica. [Quadro]
A análise dos microfilamentos de actina evidencia como um único componente molecular pode sustentar uma ampla gama de funções celulares, desde a manutenção da forma até a geração de movimento. Essa versatilidade decorre da combinação entre propriedades estruturais, dinâmica regulada e integração com sistemas motores, estabelecendo a actina como um elemento central na organização e no comportamento celular.
8.4 — Microtúbulos e organização intracelular #
A organização espacial do interior celular exige um sistema capaz de estruturar o citoplasma em múltiplas escalas, coordenando a posição de organelas, o tráfego de vesículas e a dinâmica de processos como divisão celular e polarização. Enquanto os microfilamentos de actina atuam predominantemente na interface cortical e na modulação da forma celular, os microtúbulos estabelecem uma rede de longo alcance que define a arquitetura interna da célula. Essa rede funciona como um sistema de eixos estruturais e trilhos direcionais, permitindo que a célula organize seu conteúdo de maneira eficiente e adaptativa.
Os microtúbulos são estruturas cilíndricas ocas formadas por polímeros de tubulina, cuja organização confere propriedades mecânicas e dinâmicas distintas das observadas em outros componentes do citoesqueleto. Sua distribuição no citoplasma não é aleatória, mas altamente organizada, irradiando-se a partir de centros específicos e estabelecendo padrões que refletem o estado funcional da célula. Em células em interfase, os microtúbulos organizam o posicionamento de organelas e direcionam o tráfego intracelular; durante a divisão celular, reorganizam-se profundamente para formar o fuso mitótico, responsável pela segregação cromossômica. [Figura]
8.4.1 — Estrutura dos microtúbulos e tubulina #
Os microtúbulos são formados por dímeros de tubulina, compostos por duas subunidades distintas que se associam de maneira altamente específica. Esses dímeros se organizam longitudinalmente em protofilamentos, que, por sua vez, se associam lateralmente para formar um cilindro oco com diâmetro relativamente constante. Essa estrutura tubular confere elevada resistência à compressão, permitindo que os microtúbulos suportem cargas mecânicas e mantenham a integridade estrutural da célula.
A organização dos dímeros de tubulina estabelece uma polaridade intrínseca ao microtúbulo, com extremidades estruturalmente distintas que apresentam comportamentos dinâmicos diferentes. Uma extremidade tende a favorecer a adição de novas subunidades, enquanto a outra apresenta maior tendência à perda de componentes. Essa assimetria é essencial para a função dos microtúbulos, pois permite a orientação do transporte intracelular e a reorganização direcional da rede citosquelética.
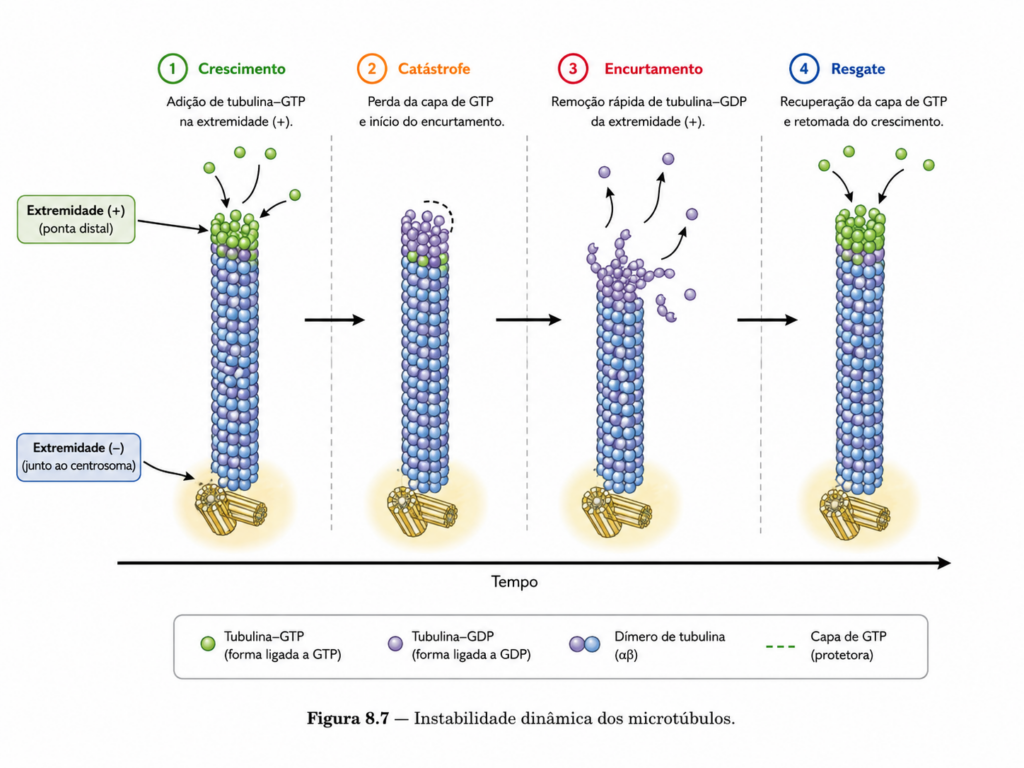
A dinâmica estrutural dos microtúbulos está diretamente associada ao estado de ligação a GTP nas subunidades de tubulina. A incorporação de dímeros associados a GTP favorece a estabilidade do filamento, enquanto a hidrólise subsequente altera a conformação da tubulina, tornando a estrutura mais suscetível à despolimerização. Esse mecanismo cria uma base molecular para o comportamento altamente dinâmico dos microtúbulos.
8.4.2 — Centros organizadores de microtúbulos (MTOCs) #
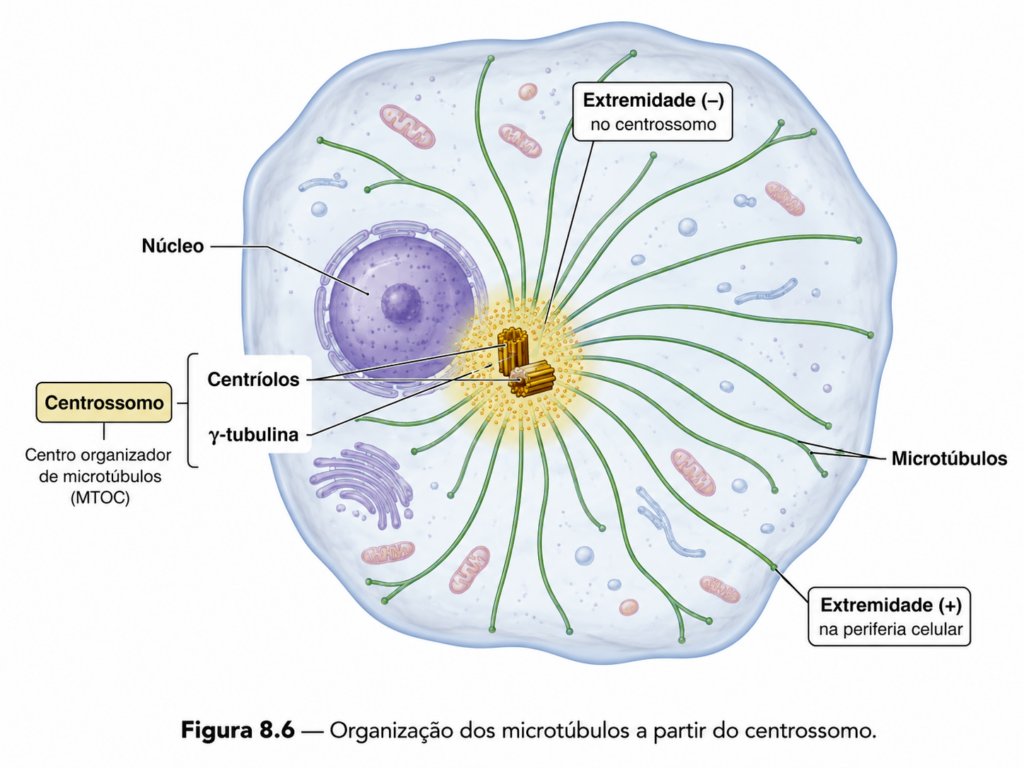
A formação e a organização dos microtúbulos dependem de regiões especializadas da célula conhecidas como centros organizadores de microtúbulos. Esses centros atuam como pontos de nucleação, onde a polimerização inicial é facilitada por complexos proteicos que estabilizam os primeiros dímeros de tubulina. A partir desses centros, os microtúbulos se estendem em direção ao citoplasma, estabelecendo uma rede radial que define a organização interna da célula.
Em células animais, o principal centro organizador é o centrossomo, que desempenha papel fundamental na organização do citoesqueleto durante a interfase e na formação do fuso mitótico durante a divisão celular. A posição do centrossomo influencia diretamente a polaridade celular e a orientação de processos como migração e divisão.
A existência de centros organizadores permite que a célula controle não apenas a formação dos microtúbulos, mas também sua orientação espacial. Esse controle é essencial para garantir que processos dependentes de transporte intracelular ocorram de maneira direcionada, evitando dispersão aleatória de organelas e vesículas.
8.4.3 — Dinâmica de instabilidade e reorganização celular #
Uma característica distintiva dos microtúbulos é seu comportamento dinâmico, conhecido como instabilidade dinâmica. Diferentemente de sistemas que apresentam crescimento contínuo ou simples equilíbrio entre montagem e desmontagem, os microtúbulos alternam entre fases de crescimento e colapso rápido. Essa alternância permite que a célula explore o espaço intracelular de maneira eficiente, ajustando rapidamente a organização de sua rede citosquelética.
Durante a fase de crescimento, a incorporação de dímeros de tubulina estabiliza a extremidade do microtúbulo, permitindo sua elongação. No entanto, a perda da “capa” de GTP pode desencadear uma transição abrupta para a fase de encurtamento, resultando em despolimerização rápida. Esse comportamento aparentemente instável é, na verdade, altamente regulado e essencial para funções celulares críticas.
No contexto da divisão celular, por exemplo, a instabilidade dinâmica permite que microtúbulos capturem cromossomos e ajustem sua posição com precisão, garantindo a correta segregação do material genético. Em células em migração, essa mesma propriedade contribui para a reorganização da rede de microtúbulos, facilitando a polarização e o direcionamento do movimento.
Além disso, a dinâmica dos microtúbulos está intimamente associada à ação de proteínas reguladoras que modulam sua estabilidade, promovendo tanto o crescimento quanto a desestabilização conforme necessário. Esse controle permite que a célula responda rapidamente a estímulos externos, reorganizando sua estrutura interna em função de mudanças no ambiente.
Em sistemas vegetais, os microtúbulos desempenham papel central na orientação da deposição de microfibrilas de celulose na parede celular, influenciando diretamente o padrão de crescimento e a forma das células. Alterações na organização dos microtúbulos podem resultar em mudanças na direção do crescimento, impactando processos de desenvolvimento e adaptação a condições ambientais adversas. [Quadro]
A análise dos microtúbulos evidencia como a combinação entre estrutura tubular, polaridade e dinâmica regulada permite a construção de um sistema altamente eficiente de organização intracelular. Essa rede não apenas sustenta a arquitetura celular, mas coordena processos complexos que dependem de orientação espacial e temporal precisa, consolidando os microtúbulos como elementos centrais na integração estrutural e funcional da célula.
8.5 — Filamentos intermediários e resistência mecânica #
A manutenção da integridade estrutural celular em ambientes sujeitos a tensões mecânicas contínuas exige a presença de elementos capazes de resistir à deformação sem comprometer a funcionalidade interna da célula. Enquanto microfilamentos e microtúbulos oferecem dinâmica e organização espacial, existe a necessidade de um componente estrutural que forneça estabilidade mecânica duradoura. Os filamentos intermediários atendem a essa demanda, constituindo um sistema especializado na resistência a forças de tração e na preservação da coesão celular.
Diferentemente de outros componentes do citoesqueleto, os filamentos intermediários não participam diretamente de processos de movimentação ativa ou transporte intracelular. Sua principal função está associada à absorção e dissipação de tensões mecânicas, protegendo a célula contra deformações excessivas que poderiam comprometer a integridade de suas estruturas internas. Essa função torna-se particularmente evidente em tecidos sujeitos a estresse físico, como epitélios, músculos e tecidos conjuntivos.
A distribuição dos filamentos intermediários no citoplasma não segue o padrão radial observado nos microtúbulos nem a organização cortical típica dos microfilamentos de actina. Em vez disso, formam uma rede densa e contínua que conecta diferentes regiões da célula, incluindo a membrana plasmática e o núcleo, estabelecendo um sistema de suporte integrado que reforça a arquitetura celular como um todo. [Figura]
8.5.1 — Diversidade estrutural dos filamentos intermediários #
Os filamentos intermediários apresentam uma característica marcante: sua diversidade molecular. Diferentes tipos celulares expressam diferentes classes de proteínas que formam esses filamentos, refletindo as demandas funcionais específicas de cada tecido. Essa diversidade não é aleatória, mas adaptativa, permitindo que cada célula possua propriedades mecânicas adequadas ao seu contexto fisiológico.
As proteínas que compõem os filamentos intermediários compartilham um padrão estrutural comum, baseado em domínios que favorecem a formação de estruturas alongadas e altamente estáveis. Essas proteínas se associam inicialmente em dímeros, que posteriormente formam tetrâmeros e, em etapas subsequentes, se organizam em filamentos robustos. Esse processo de montagem resulta em estruturas altamente resistentes, com capacidade de suportar deformações significativas sem ruptura.
[Figura]
Ao contrário de actina e tubulina, a formação dos filamentos intermediários não depende diretamente de nucleotídeos como ATP ou GTP, o que confere a essas estruturas maior estabilidade e menor dinâmica. Essa característica reforça seu papel estrutural, garantindo que a rede formada permaneça funcional mesmo em condições de estresse prolongado.
8.5.2 — Função mecânica e estabilidade celular #
A principal contribuição dos filamentos intermediários para a célula reside em sua capacidade de conferir resistência mecânica. Esses filamentos atuam como elementos de reforço que distribuem forças ao longo da célula, evitando concentrações de tensão que poderiam levar à ruptura de estruturas celulares.
Em tecidos epiteliais, por exemplo, os filamentos intermediários formam redes que conectam células adjacentes por meio de complexos de adesão, criando uma malha contínua que aumenta a resistência do tecido como um todo. Essa organização é essencial para manter a integridade de superfícies que estão constantemente expostas a forças externas, como pele e revestimentos internos de órgãos.
[Figura]
Além da resistência à tração, os filamentos intermediários também desempenham papel importante na estabilização de organelas, particularmente do núcleo. A associação desses filamentos com a lâmina nuclear contribui para a manutenção da forma nuclear e para a organização da cromatina, estabelecendo uma conexão direta entre estrutura citoplasmática e organização genética.
Em contextos patológicos, alterações na composição ou organização dos filamentos intermediários estão associadas a diversas doenças, especialmente aquelas que envolvem fragilidade tecidual. Essas condições evidenciam a importância desses filamentos na manutenção da integridade celular e na prevenção de danos estruturais.
8.5.3 — Integração com junções celulares #
A funcionalidade dos filamentos intermediários se amplia quando considerada sua integração com sistemas de adesão celular. Esses filamentos se conectam a complexos proteicos localizados na membrana plasmática, formando estruturas que ligam o citoesqueleto ao ambiente extracelular e às células vizinhas. Essa integração permite a transmissão de forças mecânicas através de múltiplas células, promovendo coesão e estabilidade em nível tecidual.
[Figura]
Entre os principais pontos de ancoragem dos filamentos intermediários estão as junções especializadas que conectam células entre si ou à matriz extracelular. Essas junções funcionam como pontos de transferência de tensão, permitindo que forças aplicadas em uma região do tecido sejam distribuídas ao longo de toda a estrutura. Esse mecanismo é essencial para evitar danos localizados e garantir a resistência global do tecido.
A interação entre filamentos intermediários e junções celulares também desempenha papel relevante na sinalização celular. A transmissão de forças mecânicas pode influenciar vias de sinalização intracelular, modulando respostas relacionadas à proliferação, diferenciação e adaptação ao ambiente. Dessa forma, os filamentos intermediários não apenas sustentam a estrutura celular, mas participam ativamente da integração entre estímulos mecânicos e respostas biológicas.
Em sistemas vegetais, embora os filamentos intermediários clássicos não estejam presentes da mesma forma que em células animais, funções análogas de resistência mecânica são desempenhadas por componentes da parede celular e por interações entre o citoesqueleto e a matriz extracelular vegetal. Essa comparação reforça a ideia de que a necessidade de suporte estrutural é uma característica universal dos sistemas biológicos, ainda que implementada por mecanismos distintos. [Quadro]
A análise dos filamentos intermediários evidencia sua importância como elementos de estabilidade em um sistema celular predominantemente dinâmico. Ao fornecer resistência mecânica e integrar diferentes regiões da célula e do tecido, esses filamentos completam o conjunto funcional do citoesqueleto, permitindo que a célula opere de maneira eficiente mesmo sob condições de estresse estrutural.
8.6 — Proteínas motoras e transporte intracelular #
A organização estrutural estabelecida pelos filamentos do citoesqueleto cria uma arquitetura capaz de sustentar e orientar a célula, mas essa organização, por si só, não explica a eficiência com que componentes celulares são distribuídos no espaço intracelular. Em um ambiente densamente ocupado, onde a difusão passiva seria insuficiente para garantir rapidez e precisão, a célula depende de mecanismos ativos de transporte. Nesse contexto, as proteínas motoras emergem como elementos centrais, convertendo energia química em movimento direcionado e transformando o citoesqueleto em uma rede funcional de trilhos dinâmicos.
Essas proteínas estabelecem a conexão entre estrutura e movimento, permitindo que vesículas, organelas e complexos macromoleculares sejam transportados de forma específica dentro da célula. Esse transporte não é aleatório, mas altamente regulado, obedecendo à polaridade dos filamentos e às necessidades fisiológicas da célula. O resultado é um sistema logístico intracelular eficiente, capaz de sustentar processos como secreção, endocitose, distribuição de organelas e manutenção da polaridade celular. [Figura]
8.6.1 — Miosinas e transporte associado à actina #
As miosinas constituem uma família de proteínas motoras que interagem com os filamentos de actina, promovendo deslocamento ao longo desses trilhos estruturais. Essas proteínas possuem domínios especializados que permitem tanto a ligação à actina quanto a hidrólise de ATP, processo que fornece a energia necessária para o movimento.
O mecanismo de ação das miosinas envolve ciclos repetitivos de ligação, mudança conformacional e deslocamento ao longo do filamento. Cada ciclo resulta em um pequeno passo, mas a repetição coordenada desses eventos gera deslocamentos significativos, permitindo o transporte de cargas intracelulares ou a geração de força mecânica.
[Figura]
Além do transporte, as miosinas desempenham papel fundamental na contração celular, especialmente em contextos onde a reorganização estrutural depende de forças internas. Em células não musculares, esse sistema contribui para processos como contração cortical, formação de anéis contráteis durante a divisão celular e ajustes na forma celular. A atuação das miosinas, portanto, vai além do transporte, integrando-se diretamente à regulação da morfologia e da dinâmica celular.
8.6.2 — Cinesinas e dineínas nos microtúbulos #
Enquanto as miosinas operam predominantemente sobre filamentos de actina, as cinesinas e dineínas são especializadas no transporte ao longo dos microtúbulos. Essas proteínas motoras utilizam a polaridade dos microtúbulos para direcionar o movimento, permitindo o transporte bidirecional dentro da célula.
As cinesinas, em geral, deslocam-se em direção a uma das extremidades do microtúbulo, promovendo o transporte de vesículas e organelas para regiões periféricas da célula. Já as dineínas realizam o movimento na direção oposta, frequentemente associadas ao transporte em direção à região central da célula. Essa organização bidirecional permite um fluxo contínuo e equilibrado de materiais, essencial para a manutenção da homeostase celular.
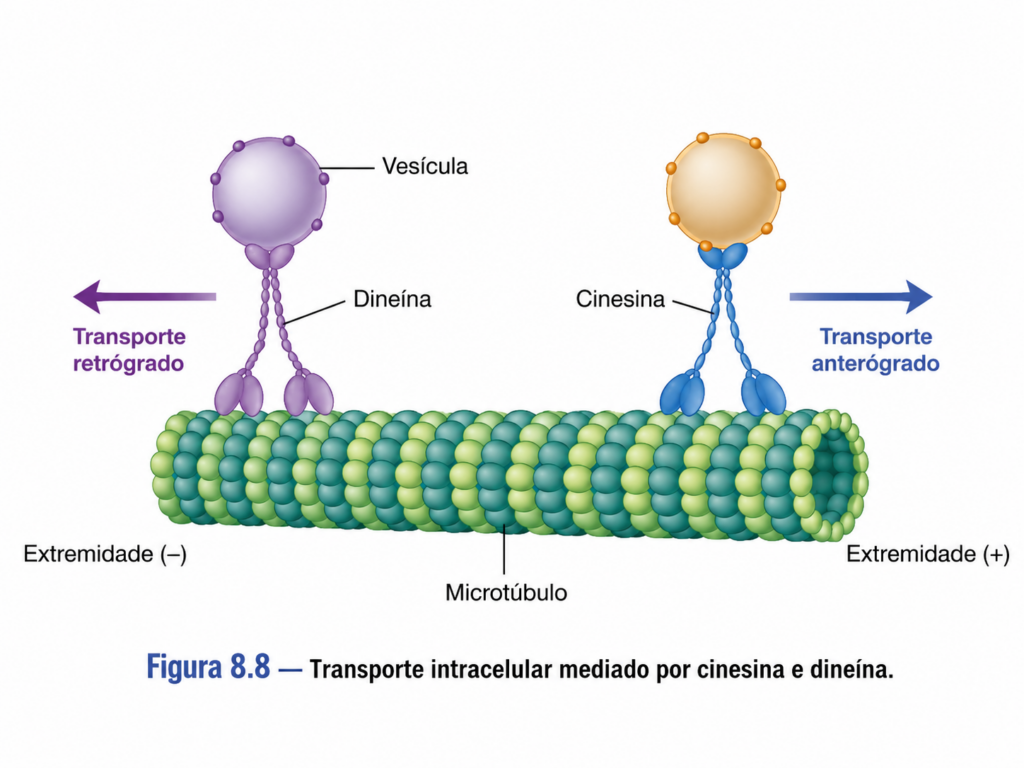
O transporte mediado por essas proteínas é altamente seletivo. Complexos proteicos específicos reconhecem e ligam cargas determinadas, garantindo que cada componente celular seja direcionado ao seu destino correto. Esse nível de especificidade é fundamental em células altamente polarizadas, como neurônios, onde longas distâncias intracelulares exigem sistemas de transporte extremamente eficientes.
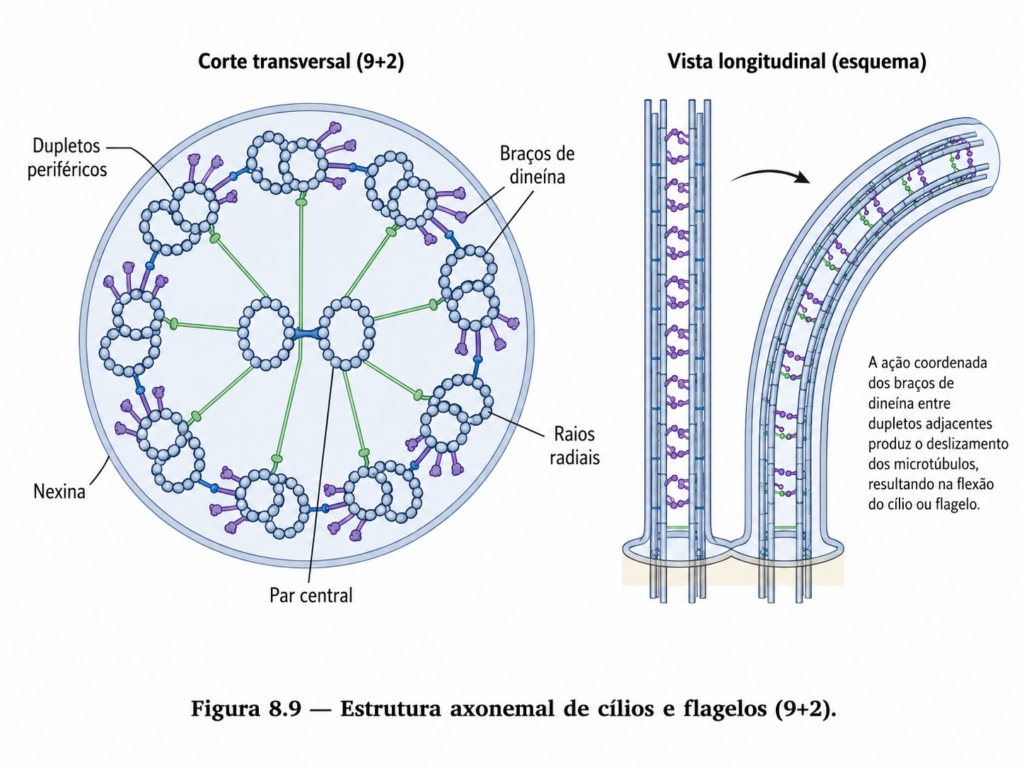
Além do transporte de cargas, as dineínas desempenham papel essencial no movimento de estruturas como cílios e flagelos, onde a interação coordenada com microtúbulos gera movimentos oscilatórios que permitem deslocamento celular ou movimentação de fluidos ao redor da célula.
8.6.3 — Conversão de energia química em movimento #
A base funcional das proteínas motoras reside na capacidade de converter energia química, proveniente da hidrólise de ATP, em trabalho mecânico. Esse processo envolve mudanças conformacionais nas proteínas motoras que são acopladas ao deslocamento ao longo dos filamentos do citoesqueleto.
Cada ciclo de hidrólise de ATP resulta em uma sequência de eventos estruturais que incluem ligação ao filamento, geração de força e liberação para um novo ciclo. Esse mecanismo garante que o movimento seja direcional e controlado, evitando deslocamentos aleatórios e permitindo a coordenação com outros processos celulares.
A eficiência desse sistema é notável, considerando a escala molecular em que opera. As proteínas motoras são capazes de transportar cargas com alta precisão, mesmo em um ambiente altamente viscoso e repleto de obstáculos. Essa capacidade sustenta não apenas o transporte intracelular, mas também a organização espacial da célula e a execução de processos complexos, como divisão celular e diferenciação.
Do ponto de vista aplicado, a compreensão desses mecanismos tem implicações diretas em áreas como biotecnologia e fisiologia vegetal. Em células vegetais, por exemplo, o transporte de vesículas mediado por proteínas motoras está diretamente relacionado à deposição de componentes da parede celular, influenciando crescimento, desenvolvimento e resposta a estresses ambientais. Alterações nesses sistemas podem impactar produtividade agrícola e adaptação a condições adversas. [Quadro]
A análise das proteínas motoras revela um nível sofisticado de integração entre energia, estrutura e função. Ao transformar o citoesqueleto em uma rede ativa de transporte e geração de força, essas proteínas consolidam a célula como um sistema dinâmico, capaz de coordenar processos complexos com precisão espacial e temporal.
8.7 — Citoesqueleto e motilidade celular #
A capacidade de movimento celular representa uma das expressões mais evidentes da natureza dinâmica do citoesqueleto. Em organismos multicelulares, a motilidade não é um evento acessório, mas um processo central para funções como desenvolvimento embrionário, cicatrização, resposta imune e adaptação ao ambiente. Em sistemas unicelulares, o movimento é frequentemente a própria condição de sobrevivência, permitindo busca por nutrientes, fuga de estímulos adversos e interação com o meio. Essa diversidade de contextos converge para um princípio comum: o deslocamento celular depende da reorganização coordenada do citoesqueleto e da geração de forças internas capazes de interagir com o ambiente externo.
A motilidade celular não ocorre por um único mecanismo, mas por diferentes estratégias estruturais que envolvem a reorganização espacial e temporal de filamentos de actina e microtúbulos, associados à atividade de proteínas motoras. Em células que se deslocam sobre superfícies, o movimento resulta de ciclos coordenados de protrusão da membrana plasmática, formação de adesões com o substrato e contração da região posterior da célula. Esse processo depende fundamentalmente da polimerização de actina na região anterior, que empurra a membrana para fora, formando estruturas como lamelipódios e filopódios. [Figura]
A formação dessas protrusões não é aleatória, mas altamente regulada por sinais intracelulares que definem a direção do movimento. A polimerização de actina ocorre de maneira polarizada, concentrando-se em regiões específicas da célula, o que estabelece uma assimetria funcional necessária para o deslocamento direcionado. À medida que novas protrusões são formadas, pontos de adesão transitórios se estabelecem entre a célula e o substrato, permitindo a transmissão de forças geradas pelo citoesqueleto. Em seguida, a contração mediada por interações entre actina e miosina promove o deslocamento do corpo celular, completando o ciclo de movimento.
Esse modelo de motilidade, frequentemente descrito como migração celular, é amplamente observado em células animais e desempenha papel crítico em processos fisiológicos e patológicos. Em tecidos, a migração celular é essencial para a reorganização estrutural durante o desenvolvimento e para a resposta a lesões. Alterações nesse processo estão associadas a condições como inflamação crônica e progressão tumoral, evidenciando a relevância da regulação do citoesqueleto para a homeostase do organismo.
Além da migração sobre superfícies, o citoesqueleto também participa de formas especializadas de movimento baseadas em estruturas altamente organizadas, como cílios e flagelos. Essas estruturas são formadas por arranjos específicos de microtúbulos associados a proteínas motoras, especialmente dineínas, que geram movimentos coordenados por meio de ciclos de deslizamento entre microtúbulos adjacentes. O resultado é um padrão oscilatório ou ondulatório que permite o deslocamento celular ou a movimentação de fluidos ao redor da célula. [Figura]
Em organismos unicelulares, os flagelos são frequentemente responsáveis pela locomoção no meio líquido, permitindo deslocamentos eficientes em ambientes aquáticos. Em organismos multicelulares, os cílios desempenham funções variadas, incluindo a movimentação de partículas ao longo de superfícies epiteliais e a participação em processos de sinalização celular. A precisão estrutural dessas organelas reflete a necessidade de coordenação fina entre os componentes do citoesqueleto para a geração de movimento eficaz.
A motilidade celular também envolve integração com sinais provenientes do ambiente. Gradientes químicos, propriedades mecânicas do substrato e interações com outras células influenciam diretamente a reorganização do citoesqueleto, modulando a direção e a intensidade do movimento. Esse processo, conhecido como migração direcionada, evidencia que o citoesqueleto não apenas executa o movimento, mas também responde ativamente a informações externas, ajustando seu comportamento de acordo com as condições ambientais.
Em sistemas vegetais, embora a motilidade celular no sentido clássico seja limitada pela presença da parede celular, processos dependentes do citoesqueleto também desempenham papel relevante na dinâmica intracelular e no crescimento orientado. A movimentação de organelas e vesículas ao longo de filamentos de actina, bem como a reorganização do citoesqueleto em resposta a estímulos como luz e gravidade, influenciam diretamente o desenvolvimento e a adaptação das plantas. [Quadro]
A análise da motilidade celular evidencia que o citoesqueleto atua como um sistema integrado de geração de força, organização estrutural e resposta ao ambiente. O movimento não emerge de um único componente, mas da interação coordenada entre filamentos, proteínas motoras e sinais regulatórios, consolidando a célula como uma unidade funcional capaz de explorar, responder e se adaptar ao seu meio de maneira altamente eficiente.
8.8 — Citoesqueleto na divisão celular #
A divisão celular impõe à célula uma reorganização estrutural profunda, na qual a arquitetura previamente estável precisa ser desmontada e substituída por uma configuração transitória altamente especializada. Esse processo não se limita à duplicação do material genético, mas envolve a redistribuição precisa de cromossomos, organelas e componentes citoplasmáticos entre duas células-filhas. A execução dessa tarefa exige um sistema capaz de coordenar forças, orientar movimentos e garantir fidelidade espacial e temporal, funções que são desempenhadas de forma integrada pelo citoesqueleto.
Durante a progressão do ciclo celular, especialmente na transição da interfase para a mitose, ocorre uma reorganização coordenada dos microtúbulos e dos filamentos de actina. A rede citosquelética previamente associada à manutenção da forma celular e ao transporte intracelular é parcialmente desmantelada, permitindo a formação de novas estruturas especializadas. Entre essas estruturas, o fuso mitótico representa o elemento central responsável pela segregação cromossômica. [Figura]
O fuso mitótico é formado por microtúbulos que se organizam a partir de centros organizadores localizados em polos opostos da célula. Esses microtúbulos apresentam dinâmicas altamente reguladas, permitindo a captura e o alinhamento dos cromossomos no plano equatorial. A interação entre microtúbulos e estruturas cromossômicas específicas garante que cada cromossomo seja corretamente conectado aos polos opostos, estabelecendo a base para uma segregação equilibrada do material genético.
A precisão desse processo depende da instabilidade dinâmica dos microtúbulos, que permite ajustes contínuos na interação com os cromossomos. Microtúbulos podem crescer, retrair e reorganizar-se rapidamente, corrigindo conexões inadequadas e assegurando que a distribuição cromossômica ocorra de forma simétrica. Esse comportamento dinâmico não representa instabilidade no sentido funcional, mas um mecanismo refinado de controle que aumenta a fidelidade da divisão celular.
À medida que a célula progride para a anáfase, forças geradas pela interação entre microtúbulos e proteínas motoras promovem o deslocamento dos cromossomos em direção aos polos celulares. Esse movimento resulta tanto do encurtamento dos microtúbulos associados aos cromossomos quanto da ação de proteínas motoras que geram tração ao longo dos filamentos. O resultado é uma separação ordenada das cromátides irmãs, etapa crítica para a manutenção da estabilidade genética.
Paralelamente à segregação cromossômica, o citoesqueleto participa da reorganização do citoplasma, preparando a célula para a divisão física. Nesse contexto, os filamentos de actina assumem papel central na formação do anel contrátil, uma estrutura localizada na região equatorial da célula que promove a constrição da membrana plasmática. A interação entre actina e miosina gera forças contráteis que levam à invaginação progressiva da membrana, culminando na separação das duas células-filhas. [Figura]
Esse processo, denominado citocinese, representa a etapa final da divisão celular e depende de uma coordenação precisa entre eventos nucleares e citoplasmáticos. A posição do anel contrátil é definida por sinais associados ao fuso mitótico, evidenciando a integração entre diferentes componentes do citoesqueleto. Essa comunicação garante que a divisão do citoplasma ocorra exatamente no plano onde os cromossomos foram segregados, evitando distribuição desigual de material celular.
Além de sua função mecânica, o citoesqueleto atua como plataforma para a organização de complexos regulatórios que controlam a progressão do ciclo celular. A reorganização estrutural observada durante a mitose está intimamente associada à atividade de proteínas reguladoras que coordenam eventos como condensação cromossômica, desorganização da membrana nuclear e montagem do fuso. Essa integração reforça a ideia de que o citoesqueleto não apenas executa processos físicos, mas também participa da regulação global da divisão celular.
Em sistemas vegetais, a divisão celular apresenta particularidades relacionadas à presença da parede celular. Embora o fuso mitótico desempenhe função semelhante na segregação cromossômica, a citocinese ocorre por meio da formação de uma estrutura específica que orienta a deposição de material para a nova parede celular. Esse processo depende da reorganização dos microtúbulos e da interação com sistemas de transporte intracelular, demonstrando que, mesmo em contextos distintos, o citoesqueleto permanece como elemento central na divisão celular. [Quadro]
A análise do papel do citoesqueleto na divisão celular evidencia como sua capacidade de reorganização estrutural e geração de força é essencial para a manutenção da continuidade da vida. A divisão celular não é apenas um evento de duplicação, mas um processo altamente coordenado que integra estrutura, dinâmica e regulação, assegurando que cada célula-filha receba não apenas material genético adequado, mas também a organização necessária para sustentar suas funções.
8.9 — Integração do citoesqueleto com membranas e sinalização celular #
A organização interna da célula não pode ser compreendida como um conjunto de sistemas isolados operando de forma independente. A funcionalidade celular emerge da integração entre estruturas, fluxos de informação e respostas ao ambiente. Nesse contexto, o citoesqueleto não atua apenas como um sistema mecânico ou estrutural, mas como um elemento central na interface entre a célula e seu meio, conectando a membrana plasmática aos sistemas de sinalização intracelular. Essa integração permite que estímulos externos sejam traduzidos em reorganizações estruturais e respostas funcionais coordenadas.
A membrana plasmática representa o principal ponto de contato da célula com o ambiente, sendo responsável pela recepção de sinais químicos e físicos. No entanto, a simples detecção de um estímulo não é suficiente para gerar uma resposta efetiva; é necessário que essa informação seja transmitida e amplificada no interior celular. O citoesqueleto participa ativamente desse processo ao interagir com proteínas de membrana e complexos de adesão, estabelecendo conexões físicas que permitem a transmissão de forças e sinais. [Figura]
Essas conexões são mediadas por proteínas que ligam diretamente os filamentos citosqueléticos a domínios específicos da membrana plasmática. Essa ancoragem não apenas estabiliza a posição da célula em relação ao substrato ou a outras células, mas também permite que alterações mecânicas na membrana sejam propagadas para o interior celular. Dessa forma, o citoesqueleto funciona como um sistema de transdução mecânica, capaz de converter estímulos físicos em sinais bioquímicos.
A integração entre citoesqueleto e sinalização celular é particularmente evidente em processos que envolvem a reorganização estrutural em resposta a estímulos externos. Sinais provenientes de receptores de membrana podem desencadear cascatas de eventos que resultam na ativação ou inibição de proteínas reguladoras do citoesqueleto. Essas alterações modulam a polimerização de filamentos, a atividade de proteínas motoras e a organização espacial da rede citosquelética, permitindo ajustes rápidos na forma e no comportamento celular.
[Figura]
Esse mecanismo é fundamental para processos como migração celular, onde sinais ambientais direcionam a reorganização da actina e dos microtúbulos, estabelecendo polaridade e orientando o deslocamento. De maneira semelhante, em células aderentes, a interação com a matriz extracelular influencia a organização do citoesqueleto, regulando a adesão, a proliferação e a diferenciação celular. A célula, portanto, responde ao ambiente não apenas por meio de alterações bioquímicas, mas também por meio de mudanças estruturais coordenadas.
A capacidade do citoesqueleto de atuar como plataforma de sinalização decorre de sua organização tridimensional e da presença de proteínas que funcionam como pontos de integração entre diferentes vias regulatórias. Esses pontos de integração permitem que sinais provenientes de diferentes fontes sejam combinados e processados, resultando em respostas específicas. A reorganização do citoesqueleto, nesse sentido, não é apenas uma consequência da sinalização, mas parte integrante do processo de decisão celular.
Outro aspecto relevante dessa integração é a resposta a estímulos mecânicos. Variações na rigidez do substrato, tensão aplicada à membrana ou alterações no fluxo de fluidos podem ser detectadas pela célula e convertidas em respostas estruturais e funcionais. Esse processo, conhecido como mecanotransdução, depende da capacidade do citoesqueleto de transmitir forças e reorganizar-se em resposta a essas forças. A adaptação a diferentes condições mecânicas é essencial para a sobrevivência celular em ambientes variáveis e para a manutenção da função tecidual.
[Figura]
Em sistemas vegetais, a integração entre citoesqueleto, membrana e sinalização assume papel estratégico na resposta a estresses ambientais. Alterações na organização dos filamentos citosqueléticos estão associadas à percepção de estímulos como gravidade, luz e disponibilidade hídrica, influenciando diretamente o crescimento e a orientação do desenvolvimento. A capacidade de reorganização do citoesqueleto permite que a célula vegetal ajuste sua estrutura em função de condições externas, contribuindo para a adaptação ao ambiente. [Quadro]
A análise da integração entre citoesqueleto, membranas e sinalização celular evidencia que a célula opera como um sistema altamente coordenado, no qual estrutura e informação estão profundamente interligadas. O citoesqueleto, ao conectar a superfície celular ao interior da célula e ao participar ativamente de processos de transdução de sinais, consolida-se como um elemento central na regulação do comportamento celular, permitindo respostas rápidas, específicas e adaptativas a um ambiente em constante transformação.
8.10 — Adaptações do citoesqueleto em diferentes tipos celulares #
A organização do citoesqueleto não segue um padrão rígido e universal, mas varia de acordo com o tipo celular, o estado fisiológico e as demandas funcionais impostas pelo ambiente. Embora os princípios moleculares que regem a polimerização de actina, a dinâmica dos microtúbulos e a estabilidade dos filamentos intermediários sejam conservados, a forma como esses elementos são distribuídos e regulados difere significativamente entre células. Essa variabilidade não representa uma inconsistência, mas uma adaptação funcional que permite à célula operar de maneira eficiente em contextos específicos.
Em células animais, a organização do citoesqueleto reflete a necessidade de flexibilidade, motilidade e interação dinâmica com o ambiente. A ausência de uma parede celular rígida impõe a dependência de uma rede citosquelética altamente responsiva para manutenção da forma e resistência mecânica. Nesse contexto, microfilamentos de actina formam uma rede cortical densa que controla a tensão superficial e permite deformações rápidas da membrana plasmática. Microtúbulos organizam o espaço intracelular, direcionando o transporte de vesículas e a posição de organelas, enquanto filamentos intermediários fornecem resistência à tração, protegendo a célula contra estresses mecânicos.
Essa organização é particularmente evidente em células com alta capacidade de migração, como células do sistema imune, onde o citoesqueleto é constantemente remodelado para permitir deslocamento rápido e direcionado. A polarização celular, essencial para a motilidade, depende de uma distribuição assimétrica dos componentes citosqueléticos, estabelecendo regiões especializadas de protrusão e contração. [Figura]
Em células epiteliais, a organização do citoesqueleto está intimamente associada à adesão celular e à formação de tecidos contínuos. Filamentos intermediários conectam-se a complexos de adesão, formando redes que atravessam múltiplas células e aumentam a resistência do tecido como um todo. Microfilamentos de actina participam da formação de junções celulares e da manutenção da polaridade apical-basal, enquanto microtúbulos contribuem para o transporte direcionado de vesículas, essencial para a manutenção de domínios funcionais distintos.
Em contraste, células vegetais apresentam uma organização citosquelética adaptada à presença de uma parede celular rígida, que limita deformações e motilidade no sentido clássico. Nesses sistemas, o citoesqueleto assume papel central na organização interna e no controle do crescimento celular. Microtúbulos orientam a deposição de microfibrilas de celulose na parede celular, determinando a direção do crescimento e influenciando a forma final da célula. Alterações na orientação dos microtúbulos resultam em mudanças no padrão de expansão celular, evidenciando sua importância na morfogênese vegetal.
[Figura]
Os filamentos de actina, por sua vez, desempenham papel fundamental no transporte intracelular, especialmente na movimentação de vesículas e organelas ao longo do citoplasma. Esse transporte é essencial para a distribuição de componentes necessários ao crescimento e à manutenção celular, compensando limitações impostas pela rigidez da parede celular. Em células com crescimento polarizado, como tubos polínicos, a organização da actina é altamente direcionada, permitindo fluxo contínuo de material para regiões específicas.
Além das diferenças entre células animais e vegetais, existem adaptações específicas em células altamente especializadas. Em neurônios, por exemplo, o citoesqueleto precisa sustentar estruturas extremamente alongadas, como axônios, ao mesmo tempo em que mantém transporte eficiente ao longo de grandes distâncias. Nesse contexto, microtúbulos formam feixes longitudinais que funcionam como trilhos para o transporte de vesículas e organelas, enquanto filamentos intermediários contribuem para a estabilidade estrutural dessas extensões.
Em células musculares, a organização do citoesqueleto está diretamente relacionada à geração de força. Microfilamentos de actina e proteínas motoras organizam-se em estruturas altamente especializadas que permitem contração coordenada. Embora esse arranjo represente uma especialização extrema, ele se baseia nos mesmos princípios fundamentais observados em outras células, evidenciando a versatilidade do citoesqueleto.
[Figura]
Do ponto de vista funcional, essas variações evidenciam que o citoesqueleto não deve ser interpretado como uma estrutura fixa, mas como um sistema adaptativo que se reorganiza de acordo com as necessidades da célula. Essa capacidade de adaptação permite que diferentes tipos celulares desenvolvam propriedades específicas, mantendo, ao mesmo tempo, uma base molecular comum.
Em contextos aplicados, especialmente na biologia vegetal e na produção agrícola, compreender essas adaptações é essencial para interpretar como células respondem a condições ambientais adversas. Alterações na organização do citoesqueleto podem influenciar diretamente o crescimento, a resistência a estresses e a eficiência na utilização de recursos, estabelecendo conexões claras entre organização celular e desempenho fisiológico. [Quadro]
A análise das adaptações do citoesqueleto em diferentes tipos celulares reforça a ideia de que a diversidade funcional observada nos organismos emerge de variações na organização de sistemas comuns. Ao ajustar a distribuição, a dinâmica e a interação de seus componentes, a célula constrói soluções específicas para desafios estruturais e funcionais, consolidando o citoesqueleto como um elemento central na integração entre forma, função e ambiente.
8.11 — Integração funcional do citoesqueleto: a célula como sistema mecânico dinâmico #
A análise dos componentes do citoesqueleto revela propriedades estruturais específicas e mecanismos moleculares bem definidos, mas a compreensão plena de sua relevância biológica emerge apenas quando esses elementos são considerados de forma integrada. A célula não opera por compartimentos isolados; ao contrário, sua funcionalidade depende da coordenação contínua entre sistemas que conectam forma, movimento, organização interna e resposta ao ambiente. Nesse contexto, o citoesqueleto se estabelece como uma rede funcional unificada, capaz de integrar diferentes escalas de organização e promover comportamento celular coerente.
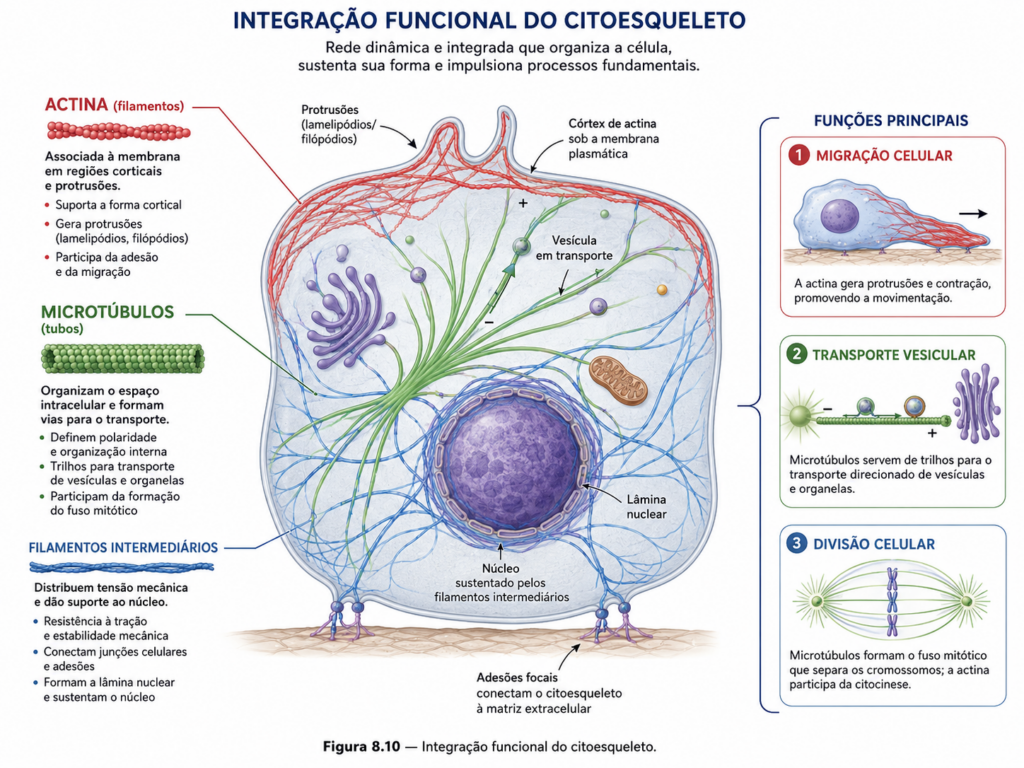
Microfilamentos de actina, microtúbulos e filamentos intermediários apresentam propriedades distintas, mas complementares. A actina atua predominantemente na interface cortical, modulando tensões e permitindo deformações rápidas da membrana plasmática. Microtúbulos estruturam o citoplasma em maior escala, organizando o posicionamento de organelas e direcionando o transporte intracelular. Filamentos intermediários, por sua vez, conferem resistência mecânica, distribuindo forças ao longo da célula e protegendo suas estruturas contra estresse físico. A integração desses três sistemas estabelece uma arquitetura que combina estabilidade e plasticidade, permitindo que a célula mantenha sua forma ao mesmo tempo em que permanece altamente adaptável. [Figura]
Essa integração é dinâmica e responsiva. Durante processos como migração celular, a polimerização de actina na região anterior é acompanhada pela reorganização dos microtúbulos, que orientam o transporte de vesículas necessárias à formação de novas adesões. Simultaneamente, filamentos intermediários redistribuem tensões, garantindo integridade estrutural durante o deslocamento. O movimento celular, portanto, não resulta da ação isolada de um único componente, mas da coordenação entre sistemas que operam em diferentes escalas e funções.
Na divisão celular, essa interdependência torna-se ainda mais evidente. A formação do fuso mitótico depende da dinâmica dos microtúbulos, enquanto a citocinese é mediada pela contração de filamentos de actina associados à miosina. A posição do anel contrátil é definida por sinais derivados do fuso, demonstrando que diferentes componentes do citoesqueleto não apenas coexistem, mas comunicam-se funcionalmente para garantir precisão na segregação do material genético e na divisão do citoplasma. [Figura]
Além da integração estrutural, o citoesqueleto atua como mediador entre sinais extracelulares e respostas intracelulares. A interação com proteínas de membrana e complexos de adesão permite que estímulos químicos e mecânicos sejam convertidos em reorganizações estruturais. Essa capacidade de transdução transforma o citoesqueleto em um elemento central na regulação do comportamento celular, influenciando processos como proliferação, diferenciação e adaptação ao ambiente.
A mecanotransdução exemplifica essa função integradora. Alterações na rigidez do substrato, na tensão aplicada à célula ou nas condições do microambiente são detectadas e propagadas através da rede citosquelética, resultando em ajustes estruturais e funcionais. Esse processo permite que a célula adapte seu comportamento a diferentes contextos, mantendo eficiência funcional mesmo em condições variáveis. [Figura]
Em sistemas vegetais, a integração entre componentes do citoesqueleto assume papel estratégico na regulação do crescimento e na resposta a estresses ambientais. A coordenação entre microtúbulos e filamentos de actina influencia a orientação da deposição de componentes da parede celular e o transporte intracelular, determinando padrões de crescimento e adaptação. Essa integração evidencia que, mesmo em contextos estruturais distintos, o citoesqueleto permanece como elemento central na organização e na funcionalidade celular. [Quadro]
A visão integrada do citoesqueleto permite compreender a célula como um sistema mecânico dinâmico, no qual forças são geradas, transmitidas e moduladas continuamente. Estrutura e função deixam de ser categorias separadas e passam a representar dimensões interdependentes de um mesmo fenômeno biológico. Essa perspectiva estabelece uma base conceitual sólida para a compreensão de processos celulares mais complexos, nos quais a organização estrutural se articula com mecanismos metabólicos e regulatórios, consolidando o citoesqueleto como um dos pilares da organização celular.



