5.1 Organização geral da célula procarionte #
A compreensão da célula procarionte exige um deslocamento conceitual importante: trata-se de um sistema biológico que opera com elevada eficiência funcional mesmo na ausência de compartimentalização interna delimitada por membranas. Essa condição não representa uma limitação estrutural, mas sim uma estratégia organizacional distinta, moldada ao longo da evolução para maximizar rapidez metabólica, adaptabilidade ambiental e economia energética. As células procariontes constituem, assim, o modelo mais fundamental de organização celular, estabelecendo o ponto de partida para a compreensão da complexidade observada nas células eucariontes.
Do ponto de vista estrutural, a célula procarionte é delimitada por uma membrana plasmática que define o limite físico entre o meio intracelular e o ambiente externo, controlando rigorosamente a entrada e saída de substâncias. Externamente a essa membrana, na maioria das bactérias, encontra-se uma parede celular rígida, responsável por conferir forma, proteção mecânica e resistência osmótica. Essa organização básica estabelece uma interface funcional crítica com o ambiente, permitindo que essas células prosperem em uma ampla variedade de condições ecológicas [Figura].
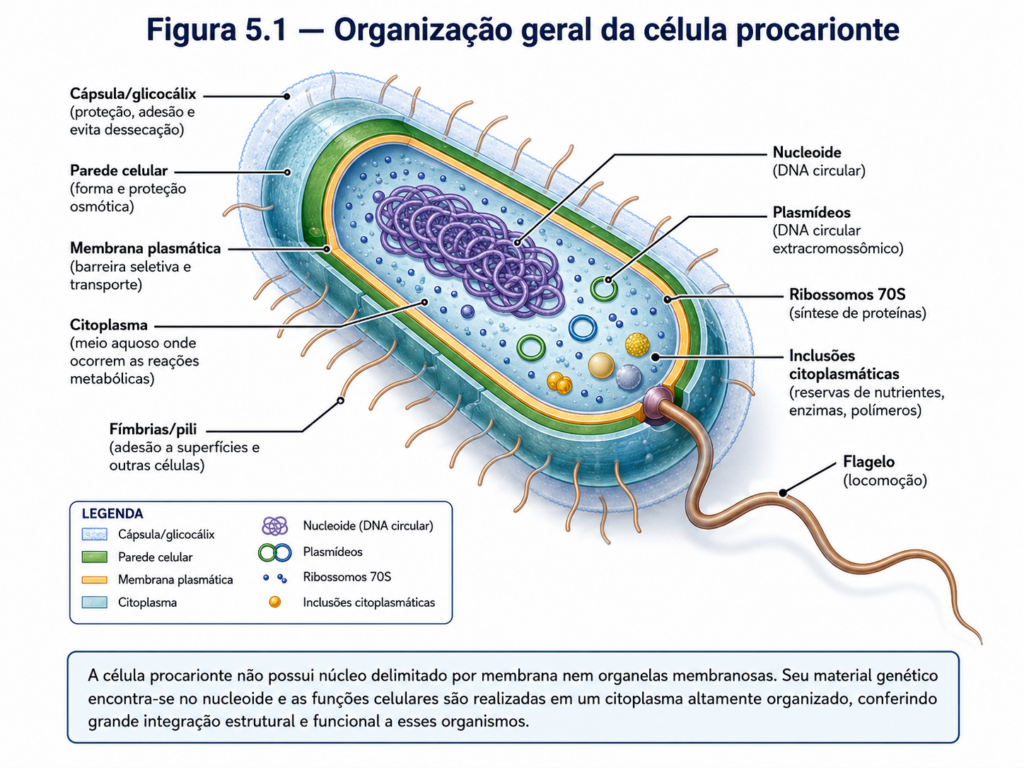
Internamente, o citoplasma apresenta uma organização aparentemente simples, mas altamente eficiente. Diferentemente das células eucariontes, não há organelas membranosas individualizadas, como núcleo, mitocôndrias ou retículo endoplasmático. No entanto, essa ausência não implica desorganização. Ao contrário, os componentes celulares estão distribuídos de maneira funcionalmente integrada, permitindo a ocorrência simultânea de múltiplos processos metabólicos. O material genético, por exemplo, encontra-se concentrado em uma região denominada nucleoide, onde o DNA, geralmente circular, permanece associado a proteínas e organizado de forma a otimizar sua replicação e expressão [Figura].
Essa organização espacial favorece uma característica central das células procariontes: a acoplamento direto entre os processos de transcrição e tradução. Como não há compartimentalização nuclear, o RNA mensageiro recém-sintetizado pode ser imediatamente acessado pelos ribossomos, permitindo uma resposta rápida às variações ambientais. Os ribossomos, por sua vez, estão dispersos no citoplasma e constituem as principais estruturas responsáveis pela síntese proteica, apresentando diferenças estruturais importantes em relação aos ribossomos eucariontes, o que tem implicações diretas em estratégias terapêuticas e biotecnológicas [Quadro].
Outro aspecto relevante da organização procarionte reside em sua escala. Essas células são, em geral, significativamente menores que as eucariontes, com dimensões típicas variando entre 1 e 10 micrômetros. Essa redução de tamanho implica uma elevada relação superfície-volume, favorecendo trocas rápidas com o meio externo e contribuindo para taxas metabólicas elevadas. Essa característica está diretamente associada à capacidade de crescimento rápido e à adaptação eficiente a mudanças ambientais, fatores que explicam a ampla distribuição dos procariontes em praticamente todos os ecossistemas do planeta [Figura].
Apesar de sua aparente simplicidade, a célula procarionte apresenta uma notável diversidade morfológica e funcional. Diferentes formas celulares, como cocos, bacilos e espirilos, refletem adaptações específicas a nichos ecológicos distintos. Além disso, estruturas especializadas, embora não universais, podem estar presentes, ampliando ainda mais as capacidades dessas células, como mecanismos de motilidade, adesão e sobrevivência em condições extremas. Essa variabilidade evidencia que a organização procarionte não deve ser interpretada como primitiva no sentido de inferioridade, mas como uma solução evolutiva altamente eficiente e versátil [Tabela].
Em termos conceituais, a distinção fundamental entre células procariontes e eucariontes reside, portanto, na forma como a informação genética e os processos metabólicos são organizados no espaço celular. Enquanto as células eucariontes utilizam compartimentos delimitados para especializar funções, as procariontes integram esses processos em um ambiente único, coordenado por interações moleculares dinâmicas. Essa diferença estrutural estabelece implicações profundas na regulação celular, na velocidade de resposta ao ambiente e na complexidade dos sistemas biológicos.
A análise da organização geral da célula procarionte revela, assim, um princípio central da biologia celular: a função emerge diretamente da estrutura, e diferentes arranjos estruturais podem sustentar estratégias igualmente eficazes de sobrevivência e adaptação. Essa compreensão fornece a base necessária para explorar, nos capítulos seguintes, a organização mais complexa das células eucariontes, permitindo estabelecer uma comparação crítica entre os dois modelos celulares e compreender os caminhos evolutivos que levaram à diversidade da vida.
5.2 Envoltório celular e interfaces com o meio #
A sobrevivência e o sucesso ecológico das células procariontes estão diretamente associados à eficiência de suas interfaces com o meio externo, uma vez que todas as trocas de matéria, energia e informação ocorrem através de estruturas que compõem o envoltório celular. Diferentemente das células eucariontes, nas quais processos metabólicos podem estar distribuídos em compartimentos internos, nos procariontes a membrana plasmática e as estruturas associadas assumem papel central na manutenção da homeostase e na mediação das interações com o ambiente. Essa condição impõe ao envoltório celular não apenas uma função de delimitação, mas também de integração funcional altamente dinâmica.
A membrana plasmática constitui o componente fundamental desse sistema, sendo formada por uma bicamada lipídica associada a proteínas que desempenham funções de transporte, reconhecimento e transdução de sinais. Sua organização segue princípios semelhantes aos observados em células eucariontes, porém com adaptações específicas que refletem a diversidade metabólica dos procariontes. Em muitas bactérias, essa membrana abriga sistemas enzimáticos essenciais, incluindo aqueles envolvidos na cadeia respiratória e na produção de ATP, evidenciando sua importância não apenas como barreira seletiva, mas como plataforma bioquímica ativa [Figura].
Externamente à membrana plasmática, a maioria das bactérias apresenta uma parede celular que confere forma e resistência mecânica, além de proteger contra variações osmóticas. Essa estrutura é predominantemente composta por peptidoglicano, uma macromolécula complexa formada por cadeias de polissacarídeos interligadas por pontes peptídicas. A organização e a espessura dessa camada variam entre diferentes grupos bacterianos, estabelecendo uma distinção clássica entre bactérias Gram-positivas e Gram-negativas. Nas Gram-positivas, a parede celular é espessa e rica em peptidoglicano, enquanto nas Gram-negativas essa camada é mais delgada e associada a uma estrutura adicional que modifica significativamente as propriedades da célula [Figura].
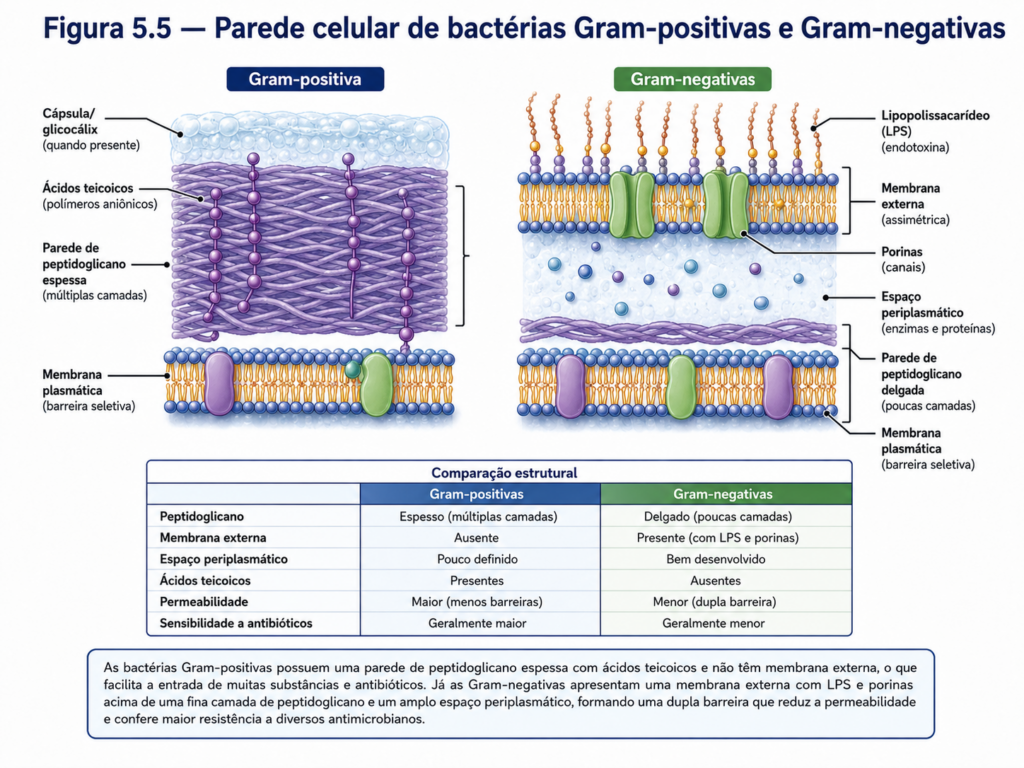
Nas bactérias Gram-negativas, a presença de uma membrana externa adiciona um nível extra de complexidade ao envoltório celular. Essa membrana, também organizada em bicamada lipídica, contém lipopolissacarídeos em sua face externa, contribuindo para a proteção contra agentes químicos e biológicos, além de participar de interações com o ambiente e com organismos hospedeiros. Entre a membrana plasmática e a membrana externa encontra-se o espaço periplasmático, uma região funcionalmente ativa que abriga enzimas, proteínas de transporte e componentes envolvidos em processos metabólicos específicos. Essa compartimentalização parcial, ainda que não equivalente à das células eucariontes, demonstra que a organização procarionte pode apresentar níveis intermediários de especialização estrutural [Figura].
Além dessas estruturas principais, muitas células procariontes apresentam camadas adicionais que ampliam sua capacidade de interação com o meio. A cápsula, composta por polissacarídeos ou proteínas, envolve a célula externamente e desempenha funções importantes na proteção contra desidratação, na evasão de mecanismos de defesa de hospedeiros e na adesão a superfícies. Essa capacidade adesiva é particularmente relevante na formação de biofilmes, comunidades microbianas organizadas que exibem comportamento coletivo e elevada resistência a condições adversas [Quadro].
Outro elemento relevante do envoltório celular são as proteínas de superfície, incluindo canais e transportadores especializados que regulam a entrada e saída de substâncias. Em bactérias Gram-negativas, proteínas como as porinas permitem a difusão controlada de pequenas moléculas através da membrana externa, enquanto sistemas de transporte mais específicos operam na membrana plasmática. Essa combinação de barreiras e mecanismos seletivos garante que a célula mantenha um ambiente interno estável, mesmo diante de variações externas significativas [Tabela].
Do ponto de vista funcional, o envoltório celular também desempenha papel central na interação com outros organismos e com o ambiente físico. Estruturas associadas, como fímbrias e pili, embora não façam parte diretamente das camadas estruturais básicas, emergem da superfície celular e contribuem para processos de adesão, colonização e transferência de material genético. Essas extensões ampliam a capacidade de resposta da célula, permitindo que ela explore nichos ecológicos específicos e estabeleça interações complexas com seu entorno [Figura].
A análise integrada do envoltório celular revela, portanto, que a interface entre a célula procarionte e o meio não é passiva, mas altamente especializada e funcionalmente diversificada. Cada componente estrutural contribui para um sistema coordenado que garante proteção, seletividade e capacidade de adaptação, elementos essenciais para a sobrevivência em ambientes frequentemente instáveis e competitivos. Essa compreensão é fundamental para interpretar não apenas a biologia dos procariontes, mas também suas aplicações em contextos médicos, agrícolas e biotecnológicos, nos quais o envoltório celular frequentemente constitui alvo estratégico de intervenções.
5.3 Componentes citoplasmáticos e organização interna #
O interior da célula procarionte costuma ser descrito, de maneira simplificada, como um espaço citoplasmático sem organelas membranosas. Essa descrição é correta, mas incompleta, porque pode induzir à ideia equivocada de que o citoplasma bacteriano corresponde a uma solução homogênea, desorganizada e meramente preenchida por moléculas dispersas. Na realidade, o citoplasma procarionte é um ambiente altamente concentrado, dinâmico e funcionalmente organizado, no qual macromoléculas, complexos enzimáticos, ribossomos, inclusões e estruturas filamentosas interagem de modo coordenado para sustentar crescimento, metabolismo, resposta ambiental e divisão celular [Figura].
A matriz citoplasmática é constituída predominantemente por água, íons, metabólitos, proteínas, ácidos nucleicos e diferentes classes de RNA. Entretanto, essa composição não deve ser interpretada como simples mistura molecular. O citoplasma é um meio fisicamente restrito, com elevada densidade macromolecular, no qual as reações bioquímicas ocorrem em condições muito diferentes daquelas observadas em soluções diluídas de laboratório. Enzimas, substratos e reguladores encontram-se em proximidade espacial, favorecendo interações rápidas e permitindo que vias metabólicas inteiras sejam organizadas por associação funcional entre proteínas. Essa condição é particularmente relevante em células pequenas, nas quais a distância entre os diferentes componentes é reduzida e a difusão molecular pode sustentar respostas celulares extremamente rápidas [Quadro].
Entre os componentes mais evidentes do citoplasma procarionte estão os ribossomos, estruturas ribonucleoproteicas responsáveis pela síntese de proteínas. Nos procariontes, os ribossomos são do tipo 70S, formados por uma subunidade menor 30S e uma subunidade maior 50S. Embora desempenhem a mesma função geral dos ribossomos eucariontes, apresentam diferenças estruturais e moleculares suficientes para serem reconhecidos como alvos seletivos de diversos antibióticos. Essa diferença permite que determinadas drogas interfiram na tradução bacteriana com menor impacto direto sobre os ribossomos citoplasmáticos das células eucariontes, embora a toxicidade e os efeitos colaterais dependam do contexto farmacológico e da especificidade de cada molécula [Figura].
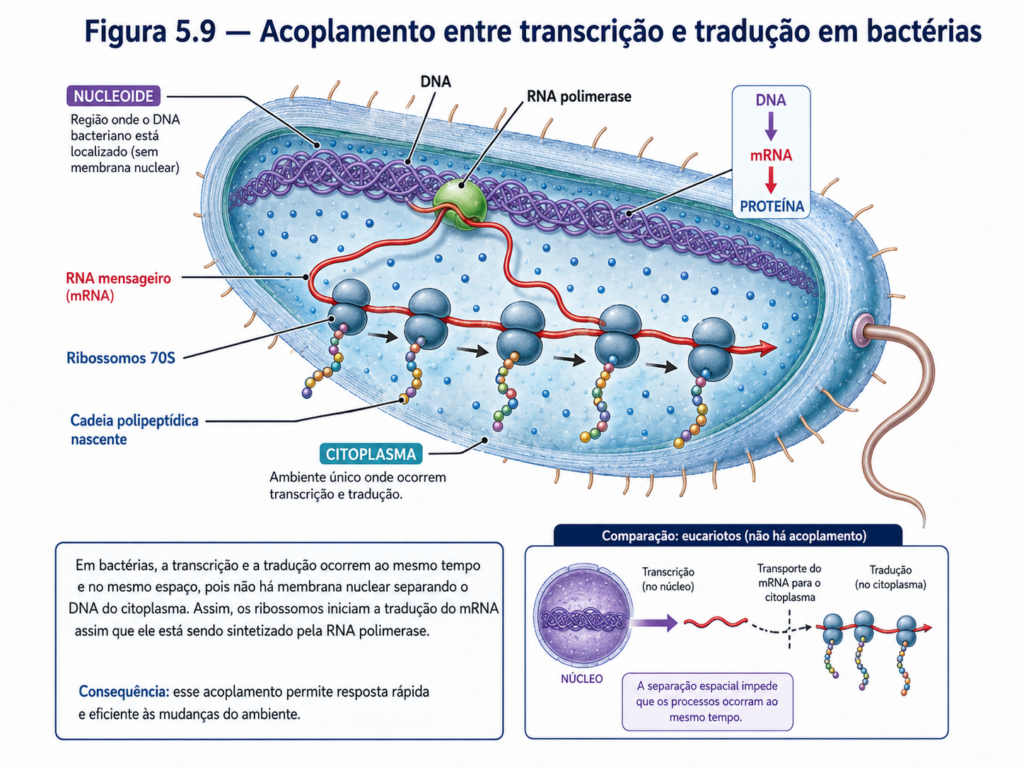
A distribuição dos ribossomos no citoplasma não é aleatória em termos funcionais. Como as células procariontes não possuem núcleo delimitado por membrana, a transcrição e a tradução podem ocorrer de maneira acoplada. Assim, enquanto uma molécula de RNA mensageiro ainda está sendo sintetizada a partir do DNA, ribossomos já podem se associar a ela e iniciar a produção da proteína correspondente. Esse acoplamento direto entre informação genética e síntese proteica representa uma das características mais marcantes da organização interna procarionte, contribuindo para a rapidez com que bactérias ajustam seu metabolismo diante de mudanças ambientais, como variações na disponibilidade de nutrientes, oxigênio, temperatura ou compostos tóxicos [Figura].
Além dos ribossomos, o citoplasma pode conter inclusões citoplasmáticas, que funcionam como depósitos temporários de substâncias ou estruturas especializadas relacionadas à sobrevivência celular. Algumas bactérias acumulam grânulos de reserva energética, como polímeros de carbono, fosfato ou enxofre, que podem ser mobilizados quando as condições ambientais se tornam desfavoráveis. Outras apresentam vesículas gasosas que contribuem para o controle de flutuabilidade em ambientes aquáticos, permitindo o posicionamento em regiões mais favoráveis à captação de luz ou nutrientes. Há ainda bactérias magnetotáticas que possuem magnetossomos, estruturas associadas a cristais minerais capazes de orientar a célula em campos magnéticos, demonstrando que a aparente simplicidade procarionte pode esconder níveis sofisticados de organização intracelular [Tabela].
Em alguns grupos bacterianos, a organização interna também inclui invaginações ou especializações da membrana plasmática associadas a processos metabólicos específicos. Em bactérias fotossintéticas, por exemplo, membranas internas derivadas ou associadas à membrana plasmática ampliam a superfície disponível para pigmentos e complexos envolvidos na captura de energia luminosa. Em outros casos, complexos proteicos ligados à membrana participam da respiração celular, do transporte de solutos e da geração de gradientes eletroquímicos. Ainda que essas estruturas não constituam organelas equivalentes às mitocôndrias ou cloroplastos, elas revelam que os procariontes podem organizar espacialmente processos bioenergéticos de modo eficiente [Figura].
A visão moderna da célula procarionte também reconhece a presença de proteínas estruturais análogas, em termos funcionais, aos componentes do citoesqueleto eucarionte. Proteínas como FtsZ, MreB e crescentina participam da divisão celular, da manutenção da forma e da organização espacial de componentes internos. FtsZ forma um anel no futuro local de divisão celular, coordenando a formação do septo durante a fissão binária. MreB está associada à manutenção da forma alongada em muitas bactérias bacilares, enquanto proteínas específicas podem contribuir para morfologias particulares. Essa descoberta modificou profundamente a interpretação clássica da célula procarionte, mostrando que sua organização interna depende de sistemas moleculares capazes de estabelecer polaridade, posicionamento e arquitetura celular [Quadro].
Essa organização estrutural é especialmente importante porque a forma celular não é apenas uma característica descritiva. Ela influencia a relação superfície-volume, a eficiência de absorção de nutrientes, a motilidade, a adesão a superfícies e a capacidade de colonizar ambientes específicos. Um bacilo, um coco ou uma bactéria espiralada não diferem apenas na aparência; essas formas refletem soluções adaptativas que resultam da interação entre parede celular, membrana, proteínas estruturais e pressões ambientais. Desse modo, a arquitetura citoplasmática e a morfologia externa devem ser interpretadas como dimensões integradas de um mesmo sistema funcional [Figura].
O citoplasma procarionte também abriga uma intensa rede de regulação molecular. Proteínas reguladoras, RNAs pequenos, metabólitos sinalizadores e complexos enzimáticos permitem que a célula ajuste rapidamente a expressão gênica e o metabolismo. Essa integração é favorecida pela ausência de barreiras internas extensas, pois sinais moleculares podem alcançar seus alvos com rapidez. Ao mesmo tempo, a célula precisa evitar interferências desordenadas entre processos simultâneos, o que exige mecanismos precisos de organização espacial e temporal. A eficiência procarionte emerge justamente desse equilíbrio entre simplicidade estrutural aparente e sofisticação molecular interna [Animação].
Assim, os componentes citoplasmáticos da célula procarionte não devem ser vistos como elementos isolados, mas como partes de uma rede funcional compacta. Ribossomos sustentam a tradução, inclusões armazenam recursos, proteínas estruturais organizam a forma e a divisão, complexos metabólicos convertem energia e moléculas reguladoras coordenam respostas ao ambiente. Essa integração permite que células pequenas, sem núcleo e sem organelas membranosas clássicas, realizem processos essenciais com grande velocidade e eficiência. A célula procarionte, portanto, não é uma célula “simples” no sentido operacional; ela é uma célula compacta, econômica e altamente otimizada para sobreviver em ambientes variáveis, competir por recursos e responder rapidamente às pressões seletivas.
5.4 Material genético e controle celular #
O material genético da célula procarionte representa um dos exemplos mais eficientes de compactação, organização e uso funcional da informação biológica. Diferentemente das células eucariontes, nas quais o DNA está contido no núcleo e associado a um sistema complexo de cromatina, os procariontes concentram seu material hereditário em uma região citoplasmática denominada nucleoide. Essa região não é delimitada por membrana, mas apresenta organização própria, resultante da associação entre DNA, proteínas estruturais, enzimas, RNAs e fatores regulatórios. Assim, a ausência de envoltório nuclear não significa ausência de controle; significa apenas que a célula utiliza outro modelo de organização da informação genética [Figura].
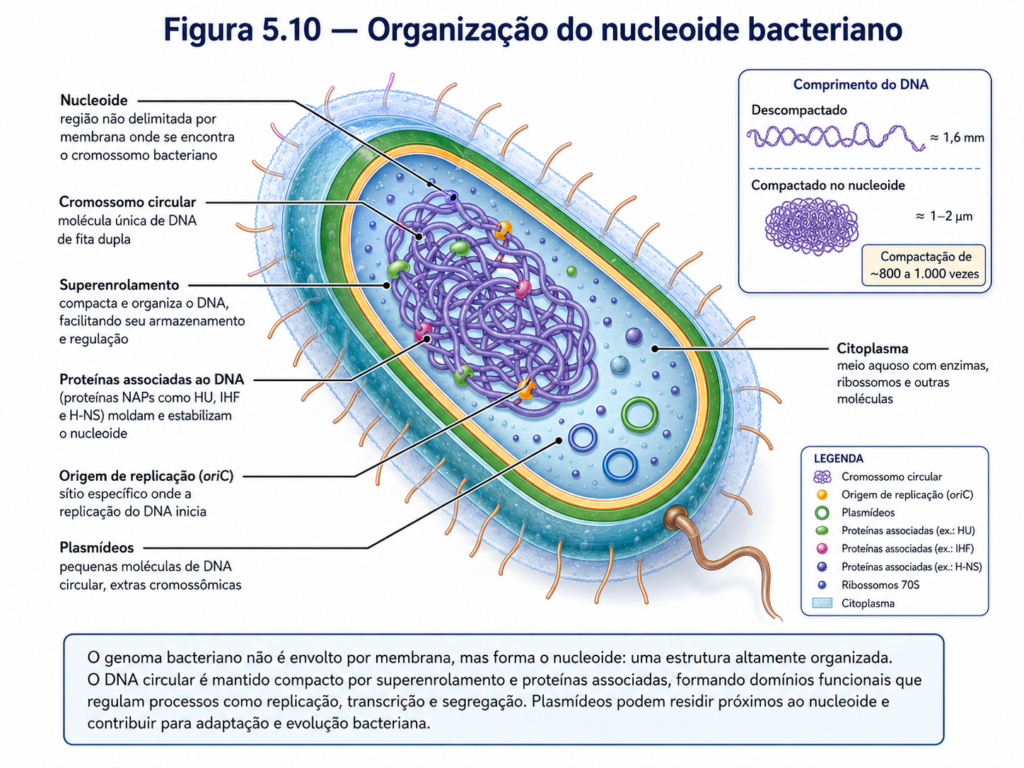
Na maioria das bactérias, o cromossomo principal é formado por uma molécula circular de DNA de dupla hélice. Essa molécula contém os genes essenciais para a manutenção da vida celular, incluindo aqueles relacionados à replicação do DNA, síntese de RNA, produção de proteínas, metabolismo energético, divisão celular e resposta ao ambiente. Como o comprimento total do DNA é muito maior que o tamanho físico da célula, ele precisa ser intensamente compactado para caber no citoplasma. Essa compactação ocorre por superenrolamento da molécula de DNA e pela ação de proteínas associadas ao nucleoide, que organizam o cromossomo em domínios funcionais [Figura].
O superenrolamento do DNA não é apenas um recurso físico de empacotamento. Ele influencia diretamente o acesso das enzimas ao material genético, a abertura da dupla hélice durante a replicação e a transcrição, e a regulação da expressão gênica. Proteínas como topoisomerases controlam o grau de tensão da molécula, permitindo que regiões específicas do cromossomo sejam copiadas, transcritas ou mantidas em estado menos acessível. Dessa forma, a arquitetura do DNA procarionte participa ativamente do controle celular, funcionando como uma dimensão regulatória integrada à própria estrutura molecular do cromossomo [Quadro].
A replicação do DNA bacteriano geralmente se inicia em uma região específica denominada origem de replicação. A partir desse ponto, a dupla hélice é aberta e duas forquilhas de replicação avançam em direções opostas ao longo do cromossomo circular, produzindo duas cópias completas do material genético. Esse processo precisa ser coordenado com o crescimento celular e com a divisão por fissão binária, de modo que cada célula-filha receba uma cópia adequada do cromossomo. Em bactérias de crescimento rápido, a replicação pode ser iniciada novamente antes que o ciclo anterior tenha sido completamente finalizado, o que contribui para taxas elevadas de proliferação em condições ambientais favoráveis [Figura].
Além do cromossomo principal, muitas bactérias possuem plasmídeos, pequenas moléculas circulares de DNA capazes de replicação independente. Embora não contenham, em geral, genes indispensáveis à sobrevivência em condições estáveis, os plasmídeos frequentemente carregam informações que conferem vantagens adaptativas em ambientes específicos. Entre essas informações estão genes de resistência a antibióticos, genes relacionados à degradação de compostos incomuns, fatores de virulência e mecanismos envolvidos na transferência genética entre células. Por isso, os plasmídeos têm grande importância ecológica, médica, agrícola e biotecnológica [Tabela].
A presença de plasmídeos ilustra uma característica central dos procariontes: a capacidade de modificar rapidamente seu repertório genético em resposta às pressões ambientais. Em populações bacterianas, genes podem circular entre células por mecanismos de transferência horizontal, permitindo que características vantajosas se disseminem com rapidez, mesmo entre linhagens não diretamente aparentadas. Esse fenômeno tem consequências profundas, como a expansão da resistência antimicrobiana, a adaptação de bactérias a ambientes contaminados e a emergência de novas capacidades metabólicas em comunidades microbianas do solo, da água, de plantas e de animais [Quadro].
O controle celular nos procariontes está fortemente associado à regulação da expressão gênica. Como essas células vivem frequentemente em ambientes sujeitos a mudanças rápidas, a capacidade de ligar ou desligar genes de acordo com a disponibilidade de nutrientes, presença de toxinas ou condições físico-químicas é essencial para sua sobrevivência. A regulação gênica bacteriana ocorre principalmente no nível da transcrição, isto é, no controle da produção de RNA a partir do DNA. Esse controle evita desperdício energético, pois a célula só sintetiza determinadas proteínas quando elas são necessárias [Figura].
Um modelo clássico de regulação gênica em procariontes é o operon, no qual genes funcionalmente relacionados são organizados em sequência e controlados por regiões regulatórias comuns. Esse arranjo permite que um conjunto de proteínas envolvidas em uma mesma via metabólica seja produzido de forma coordenada. Quando um determinado substrato está presente ou ausente no ambiente, proteínas reguladoras podem se ligar ao DNA e alterar a atividade da RNA polimerase, ativando ou reprimindo a transcrição. Esse tipo de organização mostra que, nos procariontes, o controle da informação genética é profundamente econômico e funcionalmente integrado [Figura].
A regulação gênica também pode envolver mecanismos pós-transcricionais, incluindo RNAs pequenos, alterações na estabilidade do RNA mensageiro e controle da eficiência da tradução. Esses mecanismos permitem ajustes finos na quantidade de proteínas produzidas e ampliam a capacidade de resposta da célula. Em alguns casos, sinais metabólicos internos atuam diretamente sobre proteínas reguladoras ou sobre estruturas de RNA, conectando o estado fisiológico da célula à expressão gênica. Assim, a informação genética não funciona como um programa rígido, mas como um sistema regulável, sensível ao contexto celular e ambiental [Animação].
A ausência de núcleo permite que transcrição e tradução ocorram de modo acoplado. Enquanto a RNA polimerase sintetiza uma molécula de RNA mensageiro, ribossomos podem se associar a esse RNA nascente e iniciar imediatamente a síntese proteica. Essa proximidade entre DNA, RNA e ribossomos reduz o intervalo entre sinal ambiental, resposta gênica e produção de proteínas. Em termos funcionais, isso torna a célula procarionte extremamente rápida em ajustar seu metabolismo, ativar vias de defesa, explorar novos nutrientes ou interromper processos desnecessários [Figura].
Esse modelo de organização genética tem implicações diretas para a compreensão da evolução celular. Nos procariontes, a informação hereditária não está isolada em um compartimento especializado, mas integrada ao citoplasma e ao metabolismo. O genoma, nesse contexto, é simultaneamente arquivo, centro de controle e plataforma dinâmica de resposta. Essa integração ajuda a explicar a grande plasticidade das bactérias e arqueias, sua capacidade de colonizar ambientes extremos e sua importância na história evolutiva da vida [Quadro].
Portanto, o material genético procarionte deve ser compreendido como um sistema compacto, acessível e altamente regulável. O cromossomo circular concentra as funções essenciais, os plasmídeos ampliam o repertório adaptativo, o nucleoide organiza fisicamente a informação e os mecanismos de regulação gênica ajustam a atividade celular às condições ambientais. Essa combinação de economia estrutural e flexibilidade funcional explica por que os procariontes continuam entre os organismos mais abundantes, diversos e bem-sucedidos do planeta. No contexto deste capítulo, esse subtópico também prepara o leitor para compreender, nos capítulos posteriores, como a célula eucarionte reorganizou a informação genética dentro de um núcleo delimitado, associando-a à cromatina, à compartimentalização e a formas mais complexas de controle celular.
5.5 Estruturas especializadas e adaptação funcional #
A célula procarionte não deve ser compreendida apenas a partir de sua membrana, parede celular, citoplasma e nucleoide. Embora esses componentes definam sua arquitetura básica, muitos procariontes apresentam estruturas especializadas que ampliam sua capacidade de locomoção, adesão, comunicação, transferência genética, resistência ambiental e colonização de superfícies. Essas estruturas não estão presentes em todas as espécies, nem aparecem sempre com a mesma organização, mas representam adaptações funcionais decisivas para a sobrevivência em ambientes variáveis. Em termos biológicos, elas mostram que a simplicidade estrutural procarionte não equivale à pobreza funcional; ao contrário, a célula bacteriana pode incorporar dispositivos altamente eficientes para resolver problemas específicos impostos pelo ambiente [Figura].
Entre as estruturas especializadas mais conhecidas estão os flagelos, apêndices filamentosos responsáveis pela motilidade em muitas bactérias. O flagelo bacteriano é formado por uma base inserida no envoltório celular, um gancho flexível e um filamento externo alongado, geralmente composto pela proteína flagelina. Diferentemente dos flagelos eucariontes, que se movimentam por deslizamento de microtúbulos mediado por dineínas, o flagelo bacteriano funciona como uma estrutura rotatória, impulsionada por gradientes eletroquímicos através da membrana. Essa diferença evidencia que estruturas com nomes semelhantes podem apresentar origem, composição e mecanismos completamente distintos nos dois tipos celulares [Figura].
A motilidade flagelar permite que a célula se desloque em resposta a estímulos químicos, físicos ou ambientais. Esse comportamento, conhecido como taxia, pode direcionar a bactéria em direção a substâncias favoráveis, como nutrientes, ou afastá-la de compostos tóxicos. A quimiotaxia bacteriana constitui um exemplo clássico de integração entre percepção ambiental, sinalização intracelular e resposta mecânica. A célula detecta alterações na concentração de moléculas no ambiente, processa essa informação por meio de proteínas reguladoras e ajusta o padrão de rotação dos flagelos, alternando movimentos mais direcionados com mudanças de trajetória. Essa resposta, aparentemente simples, revela um sistema de decisão celular baseado em sinais moleculares e retroalimentação [Animação].
Além dos flagelos, muitas bactérias apresentam fímbrias e pili, estruturas filamentosas mais curtas e numerosas, geralmente relacionadas à adesão a superfícies, tecidos ou outras células. As fímbrias contribuem para a fixação bacteriana em substratos específicos, o que pode ser vantajoso tanto em ambientes naturais, como partículas de solo e superfícies vegetais, quanto em interações com organismos hospedeiros. Em contextos patogênicos, essa capacidade adesiva pode representar o primeiro passo da colonização, permitindo que a bactéria permaneça no local adequado para multiplicação e formação de comunidades microbianas [Figura].
Os pili, por sua vez, podem desempenhar funções mais específicas, incluindo a conjugação bacteriana. Durante esse processo, uma célula doadora estabelece contato físico com uma célula receptora e transfere material genético, frequentemente plasmidial. Essa transferência não corresponde à reprodução sexuada no sentido eucarionte, mas constitui importante mecanismo de recombinação e disseminação de genes em populações bacterianas. Genes associados à resistência a antibióticos, fatores de virulência ou novas capacidades metabólicas podem se espalhar rapidamente por meio desse tipo de interação, aumentando a plasticidade evolutiva dos procariontes [Quadro].
A adesão mediada por fímbrias, pili, cápsulas e moléculas de superfície também está associada à formação de biofilmes. Biofilmes são comunidades microbianas organizadas, aderidas a superfícies e envolvidas por uma matriz extracelular produzida pelos próprios microrganismos. Essa matriz pode conter polissacarídeos, proteínas, lipídios, ácidos nucleicos extracelulares e água, formando uma estrutura protetora que modifica profundamente o comportamento das células envolvidas. Dentro de um biofilme, as bactérias podem apresentar metabolismo reduzido, comunicação química, maior tolerância a agentes antimicrobianos e divisão de funções entre subpopulações celulares [Figura].
A formação de biofilmes tem enorme relevância ecológica, médica, industrial e agrícola. Em ambientes naturais, biofilmes participam da ciclagem de nutrientes, da colonização de raízes, da formação de comunidades microbianas no solo e da interação entre microrganismos e superfícies minerais ou vegetais. Em sistemas agrícolas, biofilmes podem favorecer associações benéficas na rizosfera, contribuindo para proteção contra patógenos, solubilização de nutrientes ou promoção do crescimento vegetal. Em contrapartida, também podem dificultar o controle de fitopatógenos, contaminar sistemas de irrigação, obstruir tubulações e favorecer a persistência de microrganismos indesejáveis em ambientes produtivos [Quadro].
Outra estrutura especializada de grande importância adaptativa é o endósporo bacteriano. Diferentemente de uma forma reprodutiva, o endósporo representa uma estrutura de resistência, produzida por alguns grupos bacterianos quando as condições ambientais se tornam desfavoráveis. Durante a esporulação, a célula reorganiza profundamente seu conteúdo, compacta o DNA, reduz a atividade metabólica e forma camadas protetoras ao redor do material essencial. O resultado é uma estrutura altamente resistente ao calor, dessecação, radiação, agentes químicos e escassez nutricional [Figura].
A capacidade de formar endósporos permite que determinadas bactérias sobrevivam por longos períodos em estado dormente, retomando o crescimento quando as condições voltam a ser favoráveis. Esse processo é relevante para a ecologia microbiana do solo, para a conservação de populações bacterianas em ambientes extremos e para desafios sanitários, uma vez que esporos de algumas espécies são difíceis de eliminar por procedimentos convencionais. A esporulação mostra que a adaptação procarionte não depende apenas de crescimento rápido, mas também de estratégias de persistência, dormência e reativação [Tabela].
Além das estruturas de motilidade, adesão e resistência, algumas bactérias apresentam especializações relacionadas à orientação espacial e à exploração de nichos ambientais específicos. Vesículas gasosas, por exemplo, permitem que certas bactérias aquáticas regulem sua flutuabilidade e se posicionem em profundidades favoráveis à disponibilidade de luz, oxigênio ou nutrientes. Magnetossomos, presentes em bactérias magnetotáticas, contêm cristais minerais que orientam a célula em relação ao campo magnético terrestre, favorecendo deslocamentos em ambientes aquáticos estratificados. Essas estruturas demonstram que a organização procarionte pode incorporar soluções sofisticadas de navegação ambiental [Figura].
A diversidade dessas estruturas especializadas evidencia um princípio central: a adaptação funcional dos procariontes resulta da integração entre arquitetura celular, percepção ambiental e resposta fisiológica. Flagelos permitem deslocamento; fímbrias e pili favorecem adesão e transferência genética; cápsulas e biofilmes aumentam proteção e persistência; endósporos garantem sobrevivência em condições extremas; vesículas gasosas e magnetossomos refinam o posicionamento ecológico. Cada uma dessas estruturas responde a uma pressão seletiva específica, mas todas ampliam a capacidade da célula de ocupar, explorar e persistir em ambientes distintos [Tabela].
Do ponto de vista evolutivo, essas especializações ajudam a explicar por que os procariontes são tão abundantes e diversificados. A célula procarionte combina pequeno tamanho, rápido crescimento, plasticidade genética e estruturas adaptativas modulares. Essa combinação permite que populações bacterianas respondam rapidamente a mudanças ambientais e colonizem desde superfícies de raízes até organismos animais, sedimentos, águas profundas, ambientes salinos, sistemas industriais e habitats extremos. Assim, a adaptação procarionte não é produto de uma única característica, mas da soma entre organização compacta e versatilidade estrutural.
A compreensão dessas estruturas também prepara o estudante para interpretar processos que serão retomados em diferentes capítulos. A motilidade bacteriana antecipa discussões sobre citoesqueleto e movimento celular; a adesão e os biofilmes dialogam com o estudo de matriz extracelular, superfície celular e comunicação; a transferência genética se conecta à variabilidade e à evolução; a resistência por endósporos ajuda a compreender sobrevivência celular, controle microbiológico e persistência ambiental. Dessa forma, o estudo das estruturas especializadas dos procariontes revela que a célula não é apenas uma unidade delimitada por membrana, mas um sistema ativo de interação, adaptação e permanência no mundo vivo.
5.6 Metabolismo procarionte e diversidade energética #
A diversidade metabólica dos procariontes é uma das características mais importantes para compreender seu sucesso evolutivo. Enquanto os organismos eucariontes dependem, em grande parte, de vias energéticas associadas a organelas especializadas, como mitocôndrias e cloroplastos, muitos procariontes realizam processos equivalentes diretamente na membrana plasmática, em membranas internas derivadas dela ou no próprio citoplasma. Essa organização compacta permite que bactérias e arqueias explorem fontes de energia extremamente variadas, ocupando ambientes onde outros organismos não conseguem sobreviver. A célula procarionte, portanto, não deve ser vista apenas como estruturalmente simples, mas como metabolicamente versátil, capaz de converter diferentes formas de energia em trabalho celular [Figura].
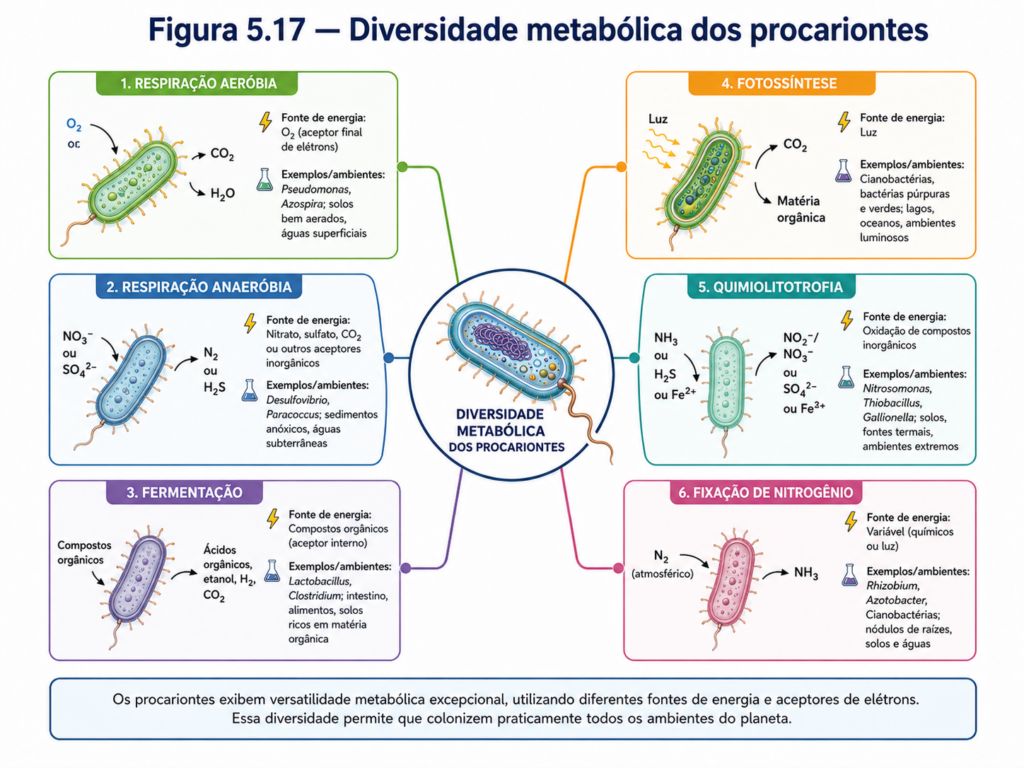
A base do metabolismo procarionte é a transformação controlada de matéria e energia. Como qualquer célula, o procarionte precisa sintetizar macromoléculas, manter gradientes iônicos, reparar danos, responder ao ambiente e se dividir. Para isso, necessita de ATP, poder redutor e precursores biossintéticos. A diferença é que os procariontes desenvolveram múltiplas formas de obter esses recursos. Alguns utilizam compostos orgânicos como fonte de carbono e energia; outros oxidam compostos inorgânicos; outros capturam energia luminosa; e alguns combinam estratégias de acordo com as condições ambientais. Essa flexibilidade metabólica amplia enormemente sua distribuição ecológica [Tabela].
Nas bactérias heterotróficas, compostos orgânicos são degradados para liberar energia e fornecer esqueletos de carbono para a biossíntese. A glicose, aminoácidos, ácidos orgânicos e lipídios podem ser usados como substratos, dependendo das enzimas disponíveis e das condições do meio. Em presença de oxigênio, muitas bactérias realizam respiração aeróbia, processo no qual moléculas orgânicas são oxidadas e os elétrons são transferidos ao oxigênio, que atua como aceptor final. A energia liberada nesse fluxo eletrônico é utilizada para gerar um gradiente eletroquímico através da membrana, o qual dirige a síntese de ATP. Nos procariontes, esse sistema está associado à membrana plasmática, que desempenha função equivalente à membrana interna mitocondrial em células eucariontes [Figura].
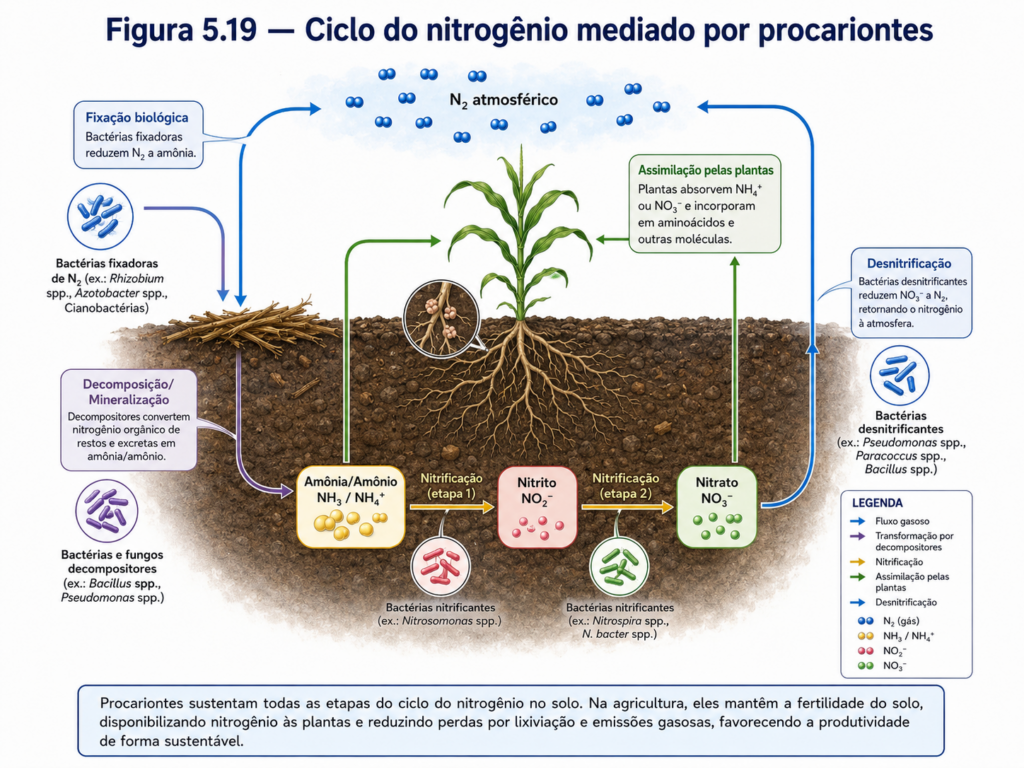
Nem todos os procariontes, porém, dependem do oxigênio. Muitos vivem em ambientes anaeróbios, como sedimentos encharcados, trato intestinal de animais, solos compactados, lodos, ambientes aquáticos estratificados ou regiões profundas pobres em oxigênio. Nesses contextos, algumas bactérias realizam respiração anaeróbia, utilizando aceptores finais de elétrons diferentes do oxigênio, como nitrato, sulfato, dióxido de carbono, ferro ou outros compostos. Esse tipo de metabolismo é essencial para ciclos biogeoquímicos, pois conecta a atividade celular microbiana à transformação de elementos como nitrogênio, enxofre, carbono e ferro no ambiente [Quadro].
A fermentação representa outra estratégia metabólica comum em procariontes. Nesse processo, a célula obtém ATP principalmente por fosforilação em nível de substrato, sem depender de uma cadeia respiratória completa ou de um aceptor externo de elétrons. Embora gere menos ATP por molécula de substrato do que a respiração aeróbia, a fermentação permite sobrevivência e crescimento em condições de baixa disponibilidade de oxigênio. Produtos fermentativos, como ácidos orgânicos, etanol, hidrogênio e dióxido de carbono, podem modificar o ambiente e servir de substrato para outros microrganismos, criando redes metabólicas cooperativas em comunidades microbianas [Figura].
Outra expressão marcante da diversidade energética procarionte é a quimiolitotrofia. Nesse modo de vida, a célula obtém energia pela oxidação de compostos inorgânicos, como amônia, nitrito, sulfeto de hidrogênio, ferro ferroso ou hidrogênio molecular. Muitos desses organismos utilizam dióxido de carbono como fonte de carbono, atuando como produtores primários em ambientes onde a luz é ausente ou limitada. Essa capacidade é fundamental em ecossistemas extremos, como fontes hidrotermais, solos minerais, ambientes ácidos e sedimentos profundos. A existência desses metabolismos mostra que a vida celular não depende exclusivamente da energia solar direta, mas pode ser sustentada por gradientes químicos geológicos [Quadro].
No contexto agrícola, a diversidade metabólica procarionte adquire relevância direta. Bactérias nitrificantes oxidam amônia a nitrito e nitrato, participando da disponibilidade de nitrogênio para as plantas. Bactérias desnitrificantes, por outro lado, reduzem nitrato em condições anaeróbias, podendo devolver nitrogênio à atmosfera e reduzir a eficiência do uso de fertilizantes nitrogenados. Outros grupos realizam fixação biológica de nitrogênio, convertendo N₂ atmosférico em formas assimiláveis, processo essencial para sistemas agrícolas sustentáveis e para a produtividade de leguminosas. Assim, o metabolismo microbiano do solo influencia diretamente fertilidade, nutrição vegetal, emissão de gases e manejo agrícola [Tabela].
A fotossíntese procarionte também apresenta grande diversidade. Cianobactérias realizam fotossíntese oxigênica, utilizando água como doadora de elétrons e liberando oxigênio como produto. Esse processo teve impacto profundo na história da Terra, pois contribuiu para a oxigenação da atmosfera e abriu caminho para a evolução da respiração aeróbia em larga escala. Outros grupos bacterianos realizam fotossíntese anoxigênica, utilizando compostos como sulfeto de hidrogênio ou moléculas orgânicas como doadores de elétrons, sem liberação de oxigênio. Essas diferentes formas de fotossíntese revelam que a captura de energia luminosa evoluiu em múltiplos contextos bioquímicos antes da consolidação dos sistemas fotossintéticos observados em plantas e algas [Figura].
Em muitos procariontes fotossintéticos, os pigmentos e complexos de transporte de elétrons estão associados à membrana plasmática ou a sistemas membranosos internos. Essa disposição reforça a importância da membrana como centro energético da célula procarionte. Enquanto nas células vegetais a fotossíntese ocorre em cloroplastos, nas cianobactérias ela depende de membranas internas especializadas, como os tilacoides bacterianos. Essa comparação é importante porque ajuda a compreender a origem evolutiva dos cloroplastos, considerados descendentes de procariontes fotossintéticos incorporados por ancestrais eucariontes [Figura].
A geração de ATP nos procariontes está frequentemente associada à força próton-motriz, criada pelo bombeamento de prótons através de membranas. O retorno desses prótons por meio da ATP sintase permite converter energia eletroquímica em energia química armazenada no ATP. Esse princípio, chamado acoplamento quimiosmótico, é compartilhado por bactérias, mitocôndrias e cloroplastos, reforçando a unidade molecular da vida e sugerindo uma origem evolutiva comum para os sistemas bioenergéticos. A diferença principal está na localização celular desses processos: nos procariontes, eles ocorrem na membrana plasmática ou em membranas associadas; nos eucariontes, em organelas especializadas [Animação].
Além da obtenção de energia, o metabolismo procarionte envolve intensa capacidade biossintética. Muitas bactérias conseguem sintetizar aminoácidos, nucleotídeos, lipídios, vitaminas, cofatores e componentes da parede celular a partir de precursores simples. Essa autonomia metabólica explica por que várias espécies conseguem crescer em meios relativamente simples, desde que disponham de fonte de carbono, nitrogênio, minerais e energia. Em outros casos, a célula depende de moléculas específicas do ambiente ou de interações com outros organismos, revelando que a autossuficiência metabólica varia amplamente entre os procariontes [Quadro].
Essa diversidade também sustenta interações ecológicas complexas. Em comunidades microbianas, o produto metabólico de uma espécie pode servir como substrato para outra, formando cadeias de dependência chamadas sintrofias. Em solos, rizosferas, sedimentos e biofilmes, diferentes microrganismos repartem funções metabólicas, estabilizam ciclos de nutrientes e respondem coletivamente às mudanças ambientais. Essa lógica é crucial para compreender sistemas agrícolas, compostagem, tratamento de efluentes, fermentações industriais e microbiomas associados a plantas e animais [Figura].
Do ponto de vista biotecnológico, a versatilidade metabólica procarionte é explorada em fermentações, produção de enzimas, biorremediação, síntese de moléculas de interesse industrial, produção de biofertilizantes e engenharia genética. Bactérias podem ser selecionadas ou modificadas para degradar contaminantes, produzir metabólitos, solubilizar fósforo, fixar nitrogênio, estimular crescimento vegetal ou atuar como agentes de biocontrole. Essa aplicação depende justamente da compreensão de como essas células capturam energia, assimilam nutrientes e regulam seu metabolismo em diferentes condições ambientais [Quadro].
A diversidade metabólica dos procariontes também impõe desafios. A mesma plasticidade que permite aplicações úteis favorece a persistência de patógenos, a resistência a estresses, a adaptação a antibióticos e a sobrevivência em ambientes hostis. Em ambientes clínicos, agrícolas ou industriais, controlar populações bacterianas exige compreender suas rotas metabólicas, seus limites fisiológicos e sua capacidade de alternar estratégias energéticas. Por isso, metabolismo procarionte não é apenas um tema bioquímico; é uma chave para interpretar ecologia, saúde, produção agrícola e tecnologia.
Assim, o metabolismo procarionte revela que a organização celular aparentemente simples pode sustentar uma variedade extraordinária de modos de vida. Respiração aeróbia, respiração anaeróbia, fermentação, quimiolitotrofia, fotossíntese oxigênica, fotossíntese anoxigênica e fixação de nitrogênio expressam diferentes soluções para o mesmo problema central: obter energia e matéria para manter a célula viva. Essa versatilidade explica a presença dos procariontes em praticamente todos os ambientes conhecidos e prepara o estudante para compreender, nos capítulos seguintes, como os sistemas bioenergéticos eucariontes preservam princípios moleculares semelhantes, mas os reorganizam em compartimentos especializados.
5.7 Reprodução e variabilidade genética #
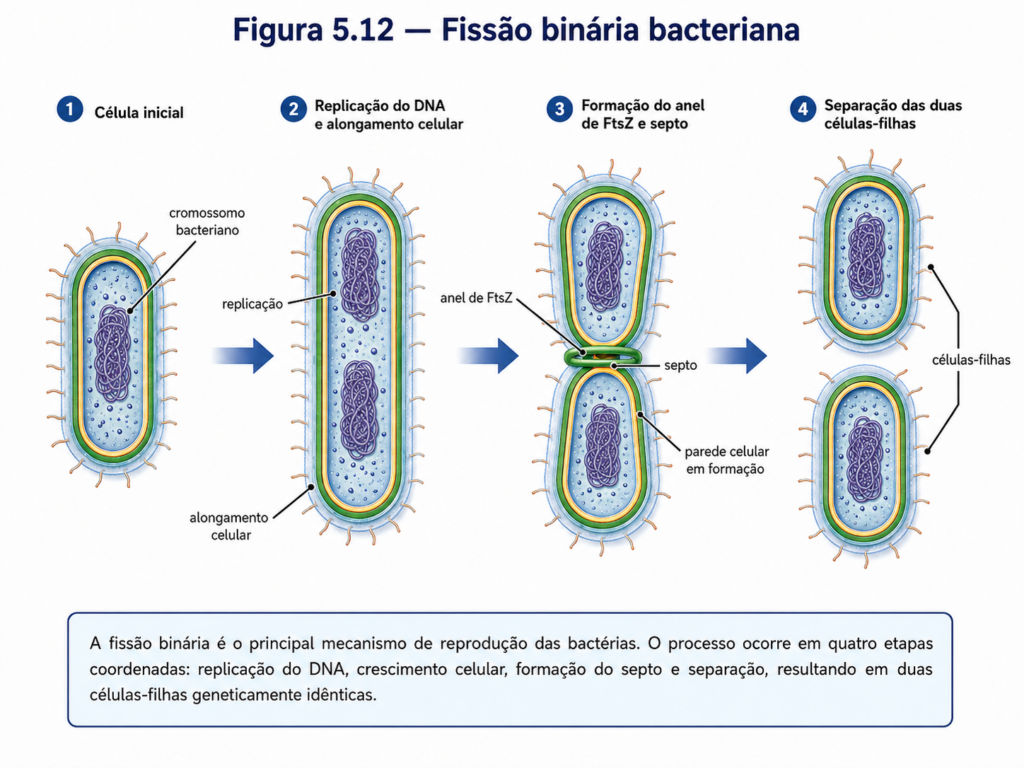
A reprodução das células procariontes está diretamente associada à simplicidade aparente de sua organização estrutural e à eficiência com que conseguem coordenar crescimento, duplicação do material genético e separação celular. Diferentemente dos eucariontes, que realizam mitose ou meiose em ciclos celulares compartimentalizados e altamente regulados, os procariontes reproduzem-se principalmente por fissão binária. Esse processo não é menos preciso por ser estruturalmente mais simples; ele representa uma solução celular compacta, rápida e funcionalmente ajustada a organismos que dependem de respostas eficientes às condições ambientais. A célula cresce, replica seu DNA, organiza a segregação dos cromossomos duplicados e divide o citoplasma em duas células-filhas geneticamente semelhantes [Figura].
Na fissão binária, o cromossomo bacteriano é duplicado a partir de uma origem de replicação, geralmente localizada em uma região específica do DNA circular. À medida que a replicação progride, as cópias cromossômicas passam a ocupar posições distintas no interior celular, em um processo coordenado com o alongamento da célula e a reorganização de proteínas estruturais. A divisão propriamente dita ocorre pela formação de um septo, uma estrutura transversal que se desenvolve no centro da célula e separa as duas futuras células-filhas. A proteína FtsZ, já apresentada como componente do citoesqueleto procarionte, forma um anel no local de divisão e recruta outras proteínas envolvidas na síntese da nova parede celular e na invaginação da membrana plasmática [Figura].
Esse mecanismo permite que muitas bactérias apresentem tempos de geração extremamente curtos quando encontram condições favoráveis de temperatura, nutrientes, pH, umidade e disponibilidade de energia. Em ambientes ideais de laboratório, algumas espécies podem duplicar sua população em intervalos de dezenas de minutos. Entretanto, essa velocidade não deve ser confundida com crescimento ilimitado. Em ambientes naturais, a proliferação bacteriana é constantemente limitada por competição, escassez de nutrientes, acúmulo de produtos tóxicos, predação por outros microrganismos, ação de bacteriófagos e variações ambientais. Assim, o crescimento populacional procarionte resulta do equilíbrio entre capacidade intrínseca de divisão e condições ecológicas concretas [Quadro].
Em cultivo fechado, como em um meio líquido de laboratório, o crescimento bacteriano costuma apresentar fases bem definidas. Inicialmente ocorre a fase lag, na qual as células ajustam seu metabolismo ao novo ambiente antes de iniciar divisão intensa. Em seguida, ocorre a fase exponencial, marcada por crescimento rápido e duplicações sucessivas. Quando nutrientes se tornam limitantes e resíduos metabólicos se acumulam, a população entra em fase estacionária, na qual o número de células que se divide tende a se equilibrar com o número de células que morre. Por fim, em condições prolongadamente desfavoráveis, instala-se a fase de declínio, caracterizada por redução progressiva da viabilidade celular [Gráfico].
A fissão binária produz, em princípio, descendentes geneticamente semelhantes à célula original. No entanto, populações procariontes não são geneticamente estáticas. A variabilidade surge por mutações espontâneas, erros de replicação, danos ao DNA, rearranjos genômicos e, sobretudo, por mecanismos de transferência horizontal de genes. Essa combinação entre reprodução rápida e geração contínua de diversidade permite que populações bacterianas explorem rapidamente novas soluções adaptativas. Em termos evolutivos, cada divisão celular aumenta o número de variantes potenciais sobre as quais a seleção natural pode atuar [Figura].
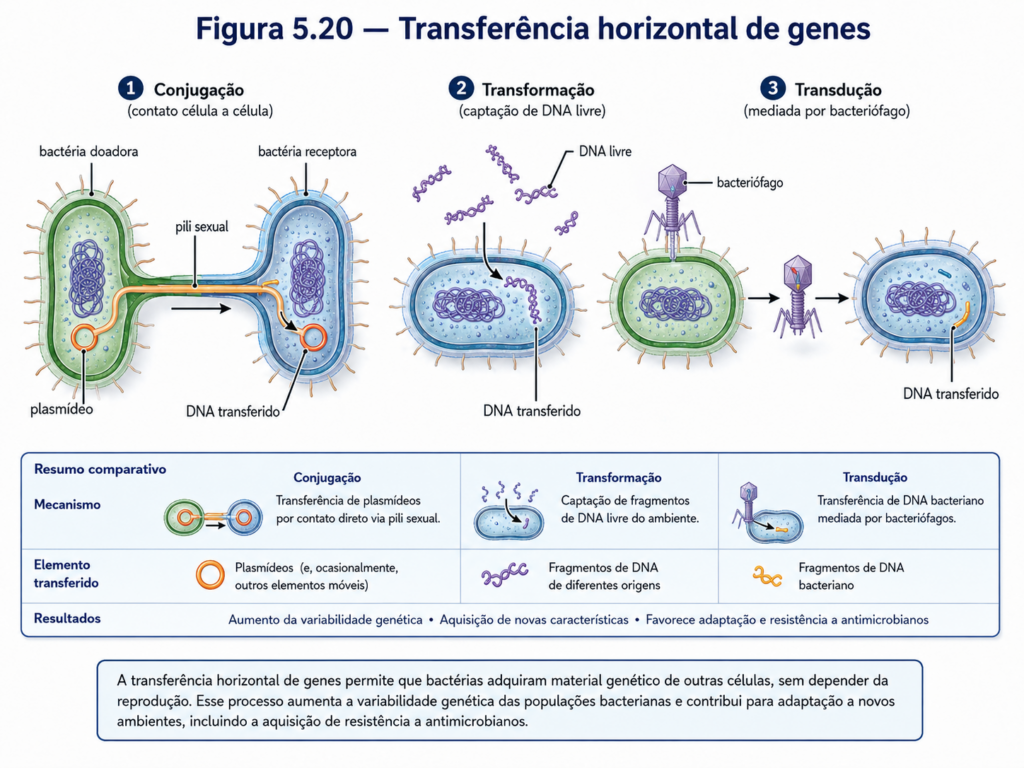
A transferência horizontal de genes constitui uma das características mais relevantes da biologia procarionte. Diferentemente da transmissão vertical, na qual a informação genética passa da célula parental para suas descendentes, a transferência horizontal permite que genes sejam adquiridos de outros organismos, inclusive de linhagens distantes. Esse processo pode alterar rapidamente propriedades funcionais de uma população, introduzindo genes relacionados à resistência a antimicrobianos, utilização de novos substratos, produção de toxinas, colonização de hospedeiros ou tolerância a condições ambientais adversas. Por isso, a evolução procarionte não pode ser compreendida apenas como uma sucessão linear de descendência, mas como uma rede dinâmica de troca genética [Quadro].
Um dos principais mecanismos de transferência horizontal é a conjugação bacteriana. Nesse processo, uma célula doadora transfere DNA para uma célula receptora por meio de contato físico direto, frequentemente mediado por pili conjugativos. O material transferido pode ser um plasmídeo ou, em alguns casos, segmentos do cromossomo bacteriano. Quando plasmídeos carregam genes adaptativos, sua disseminação por conjugação pode transformar rapidamente a composição genética de uma população. Esse mecanismo é particularmente importante na propagação de genes de resistência a antibióticos, uma vez que plasmídeos podem reunir múltiplos genes de resistência e circular entre bactérias de uma mesma comunidade [Figura].
Outro mecanismo importante é a transformação, no qual uma bactéria capta DNA livre presente no ambiente. Esse DNA pode ser proveniente de células mortas ou lisadas e, se apresentar compatibilidade molecular, pode ser incorporado ao genoma da célula receptora por recombinação. A transformação mostra que o ambiente não é apenas fonte de nutrientes e sinais, mas também reservatório potencial de informação genética. Em solos, biofilmes, sedimentos e ambientes associados a plantas, fragmentos de DNA extracelular podem persistir por períodos variáveis e contribuir para a diversidade genética microbiana [Figura].
A transdução, por sua vez, ocorre por intermédio de bacteriófagos, vírus que infectam bactérias. Durante o ciclo de infecção, fragmentos de DNA bacteriano podem ser acidentalmente incorporados a partículas virais e transferidos para outras células durante novas infecções. Esse processo transforma os fagos em vetores naturais de informação genética entre bactérias. Em alguns casos, a transdução pode disseminar genes associados à virulência, resistência ou novas capacidades metabólicas, demonstrando que vírus bacterianos têm papel importante na evolução e na ecologia das populações procariontes [Figura].
A variabilidade genética também é ampliada pela presença de elementos genéticos móveis, como transposons e integrons. Transposons são sequências capazes de mudar de posição dentro do genoma ou entre moléculas de DNA, podendo interromper genes, modificar regiões regulatórias ou transportar genes de interesse adaptativo. Integrons, por sua vez, podem capturar e expressar cassetes gênicos, funcionando como sistemas de acumulação de genes, frequentemente associados à resistência antimicrobiana. Esses elementos mostram que o genoma bacteriano é mais dinâmico do que uma simples molécula circular estável, podendo sofrer reorganizações que alteram rapidamente o fenótipo celular [Quadro].
No contexto agrícola, essa plasticidade genética tem consequências relevantes. Microrganismos do solo podem adquirir genes que ampliam sua capacidade de metabolizar compostos orgânicos, tolerar estresses ambientais, interagir com raízes ou participar de ciclos de nutrientes. A adaptação de bactérias promotoras de crescimento vegetal, simbiontes fixadores de nitrogênio e microrganismos rizosféricos depende, em parte, da variabilidade genética acumulada em populações expostas a diferentes plantas, solos, regimes hídricos e práticas de manejo. Ao mesmo tempo, fitopatógenos bacterianos também podem adquirir genes que aumentam virulência, resistência a defensivos ou capacidade de colonização, tornando o controle biológico e químico mais complexo [Quadro].
Em saúde humana e animal, a reprodução rápida e a transferência horizontal explicam parte da dificuldade de controlar infecções bacterianas. O uso inadequado ou excessivo de antibióticos cria pressão seletiva sobre populações bacterianas, favorecendo a sobrevivência de variantes resistentes. Quando genes de resistência estão localizados em plasmídeos ou elementos móveis, podem se disseminar para outras bactérias, inclusive em ambientes hospitalares, agropecuários e ambientais. Essa dinâmica evidencia que a resistência antimicrobiana não é apenas um problema clínico, mas um fenômeno ecológico e evolutivo que envolve circulação de genes entre diferentes ambientes [Tabela].
A relação entre reprodução e variabilidade genética também permite compreender por que os procariontes são tão eficientes em colonizar novos ambientes. A fissão binária aumenta rapidamente o tamanho populacional; mutações e recombinação geram diversidade; a transferência horizontal introduz genes prontos para novas funções; a seleção ambiental preserva as combinações mais vantajosas. Esse conjunto produz populações altamente adaptáveis, capazes de responder a mudanças que poderiam ser letais para organismos com ciclos reprodutivos mais lentos. Em escala evolutiva, os procariontes funcionam como sistemas de experimentação genética contínua [Figura].
É importante distinguir essa dinâmica da reprodução sexuada eucarionte. Nos procariontes, a troca genética não está obrigatoriamente vinculada à reprodução. Uma bactéria pode receber DNA por conjugação, transformação ou transdução sem que isso envolva formação de gametas, meiose ou fusão celular nos moldes eucariontes. A variabilidade genética é, portanto, parcialmente dissociada da divisão celular. Essa separação confere enorme flexibilidade evolutiva, pois a população pode se multiplicar por fissão binária e, simultaneamente, incorporar genes de diferentes fontes [Quadro].
Assim, a reprodução procarionte combina estabilidade e mudança. A fissão binária conserva a continuidade celular e permite rápida expansão populacional, enquanto os mecanismos de mutação, recombinação e transferência horizontal introduzem diversidade suficiente para adaptação. Essa lógica explica a persistência dos procariontes ao longo de bilhões de anos e sua presença em praticamente todos os ambientes conhecidos. No percurso deste capítulo, esse subtópico estabelece a ponte entre a organização estrutural da célula procarionte e sua diversidade ecológica, preparando a discussão sobre Bacteria, Archaea, extremófilos e a importância dos procariontes nos ciclos biológicos e nas aplicações biotecnológicas.
5.8 Diversidade dos procariontes #
A diversidade dos procariontes é uma das evidências mais fortes de que a complexidade biológica não depende necessariamente de grandes dimensões celulares ou de organelas membranosas. Bactérias e arqueias ocupam praticamente todos os ambientes conhecidos, desde solos agrícolas, águas superficiais e trato digestório de animais até salinas, fontes hidrotermais, geleiras, sedimentos profundos e regiões com elevada acidez ou temperatura. Essa distribuição ampla não resulta apenas de sua capacidade de reprodução rápida, mas da combinação entre plasticidade genética, versatilidade metabólica, pequeno tamanho, resistência estrutural e adaptação fisiológica. Os procariontes formam, portanto, um conjunto extremamente heterogêneo de organismos unicelulares, cuja diversidade ultrapassa em muito a imagem simplificada de “bactérias” como células pequenas e uniformes [Figura].
Durante muito tempo, os procariontes foram tratados como um grupo único, definido basicamente pela ausência de núcleo e de organelas membranosas. Essa classificação era funcional do ponto de vista didático, mas limitada do ponto de vista evolutivo. O avanço da biologia molecular, especialmente a comparação de sequências de RNA ribossômico, revelou que os organismos procariontes pertencem a dois grandes domínios da vida: Bacteria e Archaea. Ambos compartilham a ausência de núcleo delimitado por membrana, mas diferem profundamente em aspectos moleculares, bioquímicos e evolutivos. Essa distinção modificou a forma de compreender a árvore da vida e mostrou que a categoria “procarionte” descreve um tipo de organização celular, não um grupo evolutivo único e homogêneo [Figura].
As bactérias constituem o grupo procarionte mais conhecido e mais amplamente estudado. Incluem organismos de enorme importância ecológica, médica, agrícola e industrial. Algumas bactérias atuam como decompositoras, degradando matéria orgânica e devolvendo nutrientes ao ambiente. Outras participam da fixação biológica de nitrogênio, da nitrificação, da desnitrificação, da produção de compostos fermentados, da ciclagem do enxofre e da manutenção da fertilidade dos solos. Há ainda bactérias patogênicas, capazes de causar doenças em plantas, animais e seres humanos, bem como bactérias simbióticas e mutualísticas, indispensáveis ao funcionamento de microbiomas complexos [Tabela].
A diversidade bacteriana também se expressa na morfologia. Cocos, bacilos, espirilos, vibriões, filamentos e formas pleomórficas representam arranjos celulares associados a diferentes modos de vida. A forma celular influencia motilidade, adesão, resistência mecânica, capacidade de formar biofilmes, absorção de nutrientes e interação com hospedeiros. Um bacilo móvel em meio líquido, uma bactéria filamentosa em biofilme ou um coco organizado em cadeias não representam apenas variações visuais; cada organização morfológica traduz uma estratégia de ocupação do ambiente. Assim, a diversidade estrutural dos procariontes deve ser interpretada como expressão de pressões seletivas distintas [Figura].
As arqueias, por sua vez, foram inicialmente associadas a ambientes extremos, como fontes termais, salinas concentradas, lagos alcalinos, regiões anaeróbias e sedimentos profundos. Essa associação permanece importante, mas hoje se reconhece que arqueias também ocorrem em ambientes moderados, incluindo solos, oceanos e microbiomas de animais. Elas apresentam membranas com lipídios estruturalmente distintos dos bacterianos, maquinarias moleculares de transcrição e tradução mais próximas das observadas em eucariontes em certos aspectos, além de vias metabólicas particulares. Essa combinação torna as arqueias fundamentais para compreender tanto a diversidade atual da vida quanto os caminhos evolutivos que conduziram à origem das células eucariontes [Quadro].
Entre os procariontes extremófilos, a diversidade fisiológica é particularmente evidente. Termófilos vivem em temperaturas elevadas; hipertermófilos suportam condições ainda mais extremas, como as encontradas em fontes hidrotermais. Halófilos desenvolvem-se em ambientes com alta concentração de sais, exigindo adaptações específicas para manter equilíbrio osmótico e estabilidade proteica. Acidófilos prosperam em pH muito baixo, enquanto alcalófilos crescem em ambientes altamente alcalinos. Barófilos ou piezófilos toleram pressões elevadas, típicas de grandes profundidades oceânicas. Essas formas de vida demonstram que os limites da célula são mais amplos do que se imaginava quando a biologia era baseada principalmente em organismos cultivados em condições laboratoriais convencionais [Tabela].
A existência de extremófilos tem implicações científicas profundas. Em primeiro lugar, amplia o conceito de habitabilidade, mostrando que a vida pode persistir em condições consideradas hostis para plantas, animais e muitos microrganismos comuns. Em segundo lugar, contribui para hipóteses sobre a origem da vida, uma vez que alguns ambientes extremos atuais podem apresentar analogias parciais com condições da Terra primitiva. Em terceiro lugar, fornece enzimas e moléculas de interesse biotecnológico, como polimerases termoestáveis, proteínas resistentes a salinidade ou compostos capazes de funcionar em condições industriais severas. A diversidade procarionte, portanto, não é apenas um tema taxonômico; é uma fonte de modelos evolutivos e aplicações tecnológicas [Quadro].
Nos ambientes agrícolas, a diversidade dos procariontes assume papel central na produtividade e na sustentabilidade dos sistemas de cultivo. A rizosfera, região do solo influenciada pelas raízes, abriga comunidades bacterianas e arqueanas altamente dinâmicas, moldadas por exsudatos radiculares, textura do solo, umidade, disponibilidade de nutrientes, matéria orgânica e práticas de manejo. Nessa região, procariontes podem estimular o crescimento vegetal, solubilizar fósforo, produzir fitormônios, competir com patógenos, induzir resistência sistêmica em plantas ou participar da fixação de nitrogênio. Ao mesmo tempo, desequilíbrios nessas comunidades podem favorecer fitopatógenos, perda de nutrientes ou emissão de gases de efeito estufa [Figura].
A fixação biológica de nitrogênio exemplifica a importância funcional dessa diversidade. Certas bactérias livres ou simbióticas possuem a enzima nitrogenase, capaz de reduzir o nitrogênio atmosférico a formas assimiláveis. Em leguminosas, associações com rizóbios resultam na formação de nódulos radiculares, estruturas nas quais a planta fornece carbono e ambiente controlado, enquanto as bactérias contribuem com nitrogênio reduzido. Esse processo representa um dos exemplos mais relevantes de cooperação entre procariontes e organismos eucariontes, com impacto direto na agricultura, na ciclagem de nutrientes e na redução da dependência de fertilizantes nitrogenados sintéticos [Figura].
Em ambientes aquáticos, cianobactérias exercem papel ecológico igualmente decisivo. Por realizarem fotossíntese oxigênica, esses procariontes participam da produção primária e da liberação de oxigênio, além de contribuírem para ciclos do carbono e do nitrogênio. Algumas espécies fixam nitrogênio, enquanto outras podem formar florações em ambientes eutrofizados, produzindo toxinas e afetando qualidade da água, saúde animal e abastecimento humano. Esse exemplo reforça uma ideia importante: a diversidade procarionte não é automaticamente benéfica ou prejudicial; seus efeitos dependem do contexto ecológico, da composição da comunidade e das condições ambientais [Quadro].
Os procariontes também participam de interações íntimas com animais, incluindo seres humanos. Microbiomas associados ao intestino, pele, mucosas e outros ambientes corporais influenciam digestão, imunidade, metabolismo e proteção contra patógenos. Em ruminantes, comunidades microbianas anaeróbias degradam celulose e outros componentes vegetais que o animal não conseguiria digerir isoladamente. Em insetos, bactérias simbióticas podem fornecer nutrientes essenciais ou influenciar reprodução. Esses exemplos mostram que muitos organismos multicelulares funcionam, na prática, como sistemas integrados formados por células próprias e comunidades microbianas associadas [Figura].
A diversidade procarionte também se manifesta na relação com doenças. Apenas uma fração das bactérias é patogênica, mas essa fração tem grande impacto biológico e econômico. Bactérias patogênicas podem utilizar cápsulas, toxinas, sistemas de secreção, adesinas, biofilmes e mecanismos de evasão para colonizar hospedeiros e causar danos. Em plantas, fitopatógenos bacterianos podem provocar manchas foliares, murchas, podridões e perdas produtivas significativas. Em humanos e animais, infecções bacterianas continuam sendo relevantes, especialmente quando associadas à resistência antimicrobiana. Assim, compreender a diversidade bacteriana é indispensável tanto para explorar microrganismos benéficos quanto para controlar aqueles que causam prejuízos [Tabela].
Apesar da enorme diversidade conhecida, grande parte dos procariontes ainda não foi cultivada em laboratório. Durante muito tempo, o conhecimento microbiológico dependia de espécies capazes de crescer em meios artificiais, o que deixava invisível uma fração expressiva da diversidade ambiental. O uso de abordagens moleculares, como sequenciamento de DNA ambiental, metagenômica e análise de microbiomas, revelou que solos, oceanos, sedimentos, plantas e organismos animais abrigam comunidades muito mais complexas do que se imaginava. Essa mudança metodológica alterou profundamente a microbiologia e aproximou o estudo dos procariontes da ecologia molecular contemporânea [Quadro].
Essa diversidade invisível tem consequências práticas. Em solos agrícolas, por exemplo, a análise molecular pode revelar grupos microbianos associados à saúde do solo, supressão de doenças, eficiência no uso de nutrientes ou resposta a estresses hídricos. Em ambientes contaminados, pode identificar microrganismos com potencial para degradar compostos tóxicos. Em biotecnologia, pode indicar genes, enzimas e vias metabólicas ainda não exploradas. A diversidade procarionte funciona, portanto, como uma reserva biológica e genética de grande valor, cuja compreensão depende da integração entre biologia celular, genética, ecologia, bioinformática e fisiologia microbiana [Figura].
Do ponto de vista evolutivo, a diversidade dos procariontes reforça a ideia de que a vida celular não seguiu uma trajetória linear de aumento de complexidade. Bactérias e arqueias não são “formas inferiores” em relação aos eucariontes; elas representam linhagens altamente bem-sucedidas, adaptadas a estratégias celulares diferentes. A célula eucarionte ampliou a compartimentalização e a complexidade interna, enquanto os procariontes mantiveram uma organização compacta, rápida e extremamente plástica. Ambas as estratégias persistem porque resolvem problemas biológicos distintos com eficiência própria [Quadro].
Assim, a diversidade dos procariontes deve ser entendida como diversidade de formas, metabolismos, genomas, interações e estratégias ecológicas. Bacteria e Archaea compartilham um plano celular sem núcleo delimitado por membrana, mas expressam histórias evolutivas e capacidades funcionais profundamente variadas. Essa compreensão impede uma leitura simplista da célula procarionte e prepara o estudante para reconhecer sua importância nos ciclos da biosfera, nos sistemas agrícolas, na saúde, na biotecnologia e na própria evolução das células eucariontes. No conjunto do capítulo, este subtópico amplia a visão da célula procarionte: de uma estrutura celular básica para um universo de soluções biológicas que sustentam parte essencial da vida no planeta.
5.9 Procariontes no contexto aplicado: agro, saúde e biotecnologia #
A importância dos procariontes ultrapassa a descrição de sua estrutura celular. Bactérias e arqueias não são apenas modelos fundamentais para compreender a organização da vida; elas atuam diretamente nos processos que sustentam ecossistemas, sistemas produtivos, organismos multicelulares e tecnologias modernas. A mesma organização compacta que permite crescimento rápido, metabolismo versátil e adaptação genética também torna esses organismos agentes centrais em ciclos biogeoquímicos, processos infecciosos, fermentações, biorremediação, engenharia genética e inovação biotecnológica. Por isso, estudar procariontes em biologia celular não é apenas compreender células pequenas e sem núcleo; é compreender uma plataforma biológica que interfere de maneira profunda na agricultura, na saúde e na indústria [Figura].
No contexto agrícola, os procariontes são componentes essenciais da fertilidade do solo. A produtividade vegetal não depende apenas de água, luz, nutrientes minerais e manejo físico do ambiente, mas também da atividade contínua de comunidades microbianas que transformam matéria orgânica, mobilizam nutrientes e modulam interações entre raízes e solo. A rizosfera, região imediatamente influenciada pelas raízes, funciona como uma zona de intensa comunicação química, na qual exsudatos radiculares selecionam populações microbianas capazes de utilizar açúcares, aminoácidos, ácidos orgânicos e compostos fenólicos liberados pelas plantas. Em troca, diversas bactérias podem favorecer a nutrição vegetal, proteger contra patógenos ou estimular o crescimento por mecanismos diretos e indiretos [Figura].
Entre os exemplos mais relevantes está a fixação biológica de nitrogênio. O nitrogênio atmosférico é abundante, mas sua forma molecular, N₂, é quimicamente estável e não pode ser diretamente assimilada pela maioria dos organismos. Certos procariontes possuem o complexo enzimático nitrogenase, capaz de reduzir N₂ a amônia, tornando o nitrogênio biologicamente disponível. Em associações simbióticas com leguminosas, bactérias fixadoras de nitrogênio formam nódulos radiculares, nos quais recebem carbono da planta e contribuem com compostos nitrogenados. Esse processo tem implicações agronômicas profundas, pois reduz a dependência de fertilizantes nitrogenados sintéticos, diminui custos de produção e pode reduzir impactos ambientais associados à perda de nitrogênio no solo [Figura].
Além da fixação de nitrogênio, bactérias do solo participam da solubilização de fósforo, da mineralização de matéria orgânica, da produção de sideróforos, da síntese de fitormônios e da indução de resistência sistêmica em plantas. Algumas espécies produzem auxinas, giberelinas ou compostos voláteis capazes de modificar o crescimento radicular, aumentando a exploração do solo e a absorção de água e nutrientes. Outras competem com fitopatógenos por espaço e recursos, produzem antibióticos naturais ou ativam respostas defensivas da planta. Esse conjunto de funções sustenta o desenvolvimento de bioinsumos, inoculantes e agentes de biocontrole, cuja eficiência depende da compatibilidade entre microrganismo, cultura agrícola, solo, clima e manejo [Quadro].
A aplicação de procariontes na agricultura, entretanto, exige cautela técnica. Um microrganismo promissor em laboratório pode não apresentar o mesmo desempenho em campo, onde enfrenta competição com microbiotas nativas, variações de temperatura, umidade, pH, salinidade, matéria orgânica e práticas agrícolas. Por isso, bioinsumos não devem ser tratados como soluções genéricas e universais. Sua eficácia depende de formulação adequada, viabilidade celular, dose, método de aplicação, estabilidade no armazenamento e validação agronômica em condições reais. Esse ponto é decisivo para formar estudantes capazes de diferenciar biotecnologia sólida de promessa comercial mal sustentada [Tabela].
Na saúde humana e animal, os procariontes também ocupam uma posição ambígua: podem ser indispensáveis à homeostase ou agentes de doença. A microbiota associada ao intestino, pele, cavidade oral, trato respiratório e trato urogenital participa da digestão, da produção de metabólitos, da modulação imunológica e da proteção contra microrganismos patogênicos. Em ruminantes, comunidades bacterianas anaeróbias degradam celulose e outros componentes vegetais, permitindo o aproveitamento energético de materiais que o animal não degradaria sozinho. Nesses casos, o organismo multicelular funciona como um ecossistema integrado, no qual células eucariontes e procariontes mantêm relações metabólicas e imunológicas permanentes [Figura].
Por outro lado, bactérias patogênicas utilizam estruturas e mecanismos celulares para colonizar tecidos, escapar de defesas e causar danos. Cápsulas podem dificultar a fagocitose; fímbrias e adesinas favorecem a fixação a superfícies; toxinas alteram funções celulares do hospedeiro; sistemas de secreção injetam proteínas efetoras; biofilmes aumentam persistência e tolerância a tratamentos. A patogenicidade, portanto, não é uma propriedade abstrata, mas o resultado de interações moleculares específicas entre envoltório celular, metabolismo, regulação gênica, fatores de virulência e resposta do hospedeiro [Quadro].
A compreensão da organização procarionte sustenta o desenvolvimento e o uso racional de antibióticos. Muitas drogas antimicrobianas atuam sobre estruturas ou processos bacterianos que diferem dos equivalentes eucariontes, como a síntese de peptidoglicano da parede celular, a tradução nos ribossomos 70S, a replicação do DNA bacteriano ou vias metabólicas específicas. Essa seletividade explica por que é possível atingir bactérias sem destruir diretamente as células do hospedeiro. Entretanto, a seletividade nunca é absoluta e o uso inadequado desses fármacos favorece a seleção de linhagens resistentes, especialmente quando genes de resistência se disseminam por plasmídeos, transposons, integrons e transferência horizontal [Figura].
A resistência antimicrobiana representa um dos exemplos mais claros de biologia celular aplicada à realidade contemporânea. Uma população bacteriana exposta a um antibiótico não “decide” tornar-se resistente; a pressão seletiva favorece células que já possuem mutações, genes adquiridos ou mecanismos capazes de reduzir o efeito da droga. Esses mecanismos podem incluir alteração do alvo molecular, produção de enzimas inativadoras, bombas de efluxo, redução de permeabilidade da membrana ou formação de biofilmes. Assim, a resistência emerge da interação entre genética, envoltório celular, metabolismo e ecologia microbiana. Esse fenômeno conecta medicina, agropecuária, saneamento, indústria e ambiente em uma mesma problemática biológica [Tabela].
Na biotecnologia, os procariontes tornaram-se ferramentas centrais porque combinam crescimento rápido, facilidade de cultivo, genomas relativamente compactos e possibilidade de manipulação genética. A bactéria Escherichia coli, por exemplo, tornou-se um dos principais modelos experimentais da biologia molecular e da engenharia genética. Plasmídeos bacterianos são usados como vetores para clonagem de DNA, expressão de proteínas recombinantes e construção de sistemas genéticos controlados. Essa tecnologia permitiu produzir enzimas, hormônios, antígenos, proteínas industriais e moléculas de interesse científico em larga escala [Figura].
A produção de proteínas recombinantes em bactérias demonstra como uma estrutura celular aparentemente simples pode ser convertida em plataforma industrial. Um gene de interesse pode ser inserido em um plasmídeo, introduzido em uma bactéria e expresso sob controle de sequências regulatórias específicas. A célula passa então a produzir a proteína codificada, que pode ser purificada e utilizada em pesquisa, diagnóstico, indústria ou terapêutica. Esse princípio está na base de muitas tecnologias modernas, embora sua aplicação exija controle rigoroso de expressão, dobramento proteico, toxicidade celular, estabilidade plasmidial, rendimento e qualidade do produto final [Quadro].
Além da engenharia genética clássica, procariontes são utilizados em fermentações industriais. Bactérias láticas participam da produção de iogurtes, queijos, vegetais fermentados e outros alimentos; bactérias acéticas contribuem para a produção de vinagre; diferentes microrganismos são explorados na síntese de ácidos orgânicos, solventes, enzimas e biopolímeros. Nessas aplicações, o metabolismo procarionte é direcionado para gerar produtos de interesse econômico. A eficiência do processo depende da cepa utilizada, da composição do meio, do controle de temperatura, pH, oxigênio, agitação, contaminação e tempo de cultivo [Figura].
A biorremediação constitui outro campo aplicado de grande relevância. Certos procariontes são capazes de degradar hidrocarbonetos, pesticidas, solventes, compostos aromáticos, resíduos orgânicos e até participar da transformação de metais em formas menos móveis ou menos tóxicas. Essa capacidade deriva da diversidade metabólica discutida anteriormente e pode ser explorada na recuperação de solos, águas e efluentes contaminados. No entanto, a biorremediação também exige análise criteriosa, pois o sucesso depende da disponibilidade do contaminante, das condições ambientais, da presença de aceptores de elétrons, da competição microbiana e da ausência de efeitos ecológicos indesejados [Tabela].
Em biologia sintética, bactérias e arqueias são cada vez mais tratadas como chassis celulares, isto é, sistemas vivos que podem ser redesenhados para executar funções programadas. Circuitos genéticos artificiais, sensores celulares, vias metabólicas reconstruídas e microrganismos produtores de moléculas de alto valor ilustram essa tendência. A célula procarionte torna-se, nesse contexto, uma plataforma de computação biológica e produção molecular. Essa abordagem abre oportunidades para agricultura de precisão, biossensores ambientais, diagnóstico rápido, produção sustentável de insumos e novos processos industriais [Animação].
Apesar de seu potencial, o uso aplicado de procariontes exige reflexão ética, biossegurança e governança tecnológica. A liberação de microrganismos no ambiente, o uso de organismos geneticamente modificados, a disseminação de genes de resistência, a contaminação cruzada em processos industriais e os impactos sobre microbiomas naturais precisam ser avaliados com rigor. A biotecnologia microbiana não deve ser vista apenas como ferramenta produtiva, mas como intervenção sobre sistemas vivos complexos. Em ambientes agrícolas, por exemplo, introduzir um inoculante significa interferir em uma rede ecológica já existente, e não apenas adicionar uma “função” isolada ao solo [Quadro].
A integração entre agro, saúde e biotecnologia revela uma ideia central: os procariontes são simultaneamente organismos naturais, agentes ecológicos, ferramentas tecnológicas e desafios de controle. O mesmo grupo celular que fixa nitrogênio pode inspirar biofertilizantes; o mesmo princípio de transferência genética que sustenta adaptação pode disseminar resistência; a mesma rapidez de crescimento que favorece produção industrial pode gerar contaminações; a mesma capacidade de formar biofilmes pode proteger raízes ou dificultar tratamentos. Essa ambivalência exige uma leitura madura da biologia celular procarionte, evitando tanto a demonização das bactérias quanto a romantização de sua aplicação.
Assim, o estudo dos procariontes no contexto aplicado mostra que a estrutura celular tem consequências práticas. Membrana, parede, ribossomos, nucleoide, plasmídeos, metabolismo, motilidade, adesão e biofilmes não são apenas tópicos isolados de um capítulo; são fundamentos para compreender fertilidade do solo, doenças infecciosas, produção de alimentos, bioinsumos, engenharia genética, resistência antimicrobiana e biorremediação. A célula procarionte, pequena em escala microscópica, possui impacto macroscópico na biosfera e na sociedade. Essa percepção prepara o leitor para integrar os conceitos estudados até aqui e avançar para a comparação com a célula eucarionte, na qual a complexidade estrutural assume outro modelo de organização e controle.
5.10 Integração estrutural e funcional da célula procarionte #
A célula procarionte só pode ser compreendida adequadamente quando seus componentes deixam de ser vistos como partes isoladas e passam a ser interpretados como elementos de um sistema integrado. Membrana plasmática, parede celular, citoplasma, ribossomos, nucleoide, plasmídeos, inclusões, estruturas de motilidade, mecanismos de adesão e vias metabólicas não atuam separadamente. Cada um desses componentes participa de uma rede funcional compacta, na qual a sobrevivência depende da coordenação entre estrutura, informação genética, metabolismo e interação ambiental. Essa integração é o que permite que células sem núcleo e sem organelas membranosas clássicas apresentem crescimento rápido, elevada adaptabilidade e ampla distribuição ecológica [Figura].
O primeiro nível dessa integração ocorre na própria delimitação celular. A membrana plasmática não é apenas uma barreira física; ela regula fluxos de íons, nutrientes, resíduos metabólicos e sinais ambientais. Ao mesmo tempo, atua como plataforma para processos energéticos, especialmente em bactérias que realizam respiração ou fotossíntese associada a sistemas de transporte de elétrons. A parede celular, por sua vez, estabiliza a forma, protege contra variações osmóticas e condiciona a interação da célula com o meio. Assim, o envoltório celular conecta diretamente proteção, forma, transporte, metabolismo energético e comunicação com o ambiente [Figura].
Essa integração entre membrana e metabolismo é particularmente importante nos procariontes porque muitas funções que, em eucariontes, estão associadas a organelas especializadas ocorrem na própria membrana plasmática ou em suas especializações. O gradiente eletroquímico usado para síntese de ATP, por exemplo, depende da separação entre o interior celular e o meio externo. Essa diferença de concentração e carga elétrica, mantida por proteínas de membrana, converte a superfície celular em um sistema bioenergético ativo. Dessa forma, a membrana não apenas separa a célula do ambiente; ela transforma essa separação em fonte de trabalho biológico [Animação].
No interior celular, o citoplasma funciona como um espaço de reação altamente concentrado, no qual moléculas e complexos macromoleculares interagem com elevada eficiência. A ausência de compartimentos membranosos não elimina a organização interna; ela exige outra lógica de organização, baseada em proximidade molecular, domínios funcionais, associação entre proteínas e distribuição espacial de estruturas. Ribossomos, enzimas, RNAs, metabólitos e proteínas reguladoras compartilham o mesmo ambiente, permitindo que processos como transcrição, tradução, metabolismo e resposta a sinais ocorram em estreita coordenação [Figura].
O acoplamento entre transcrição e tradução é um exemplo central dessa integração. Como o DNA não está isolado por um envoltório nuclear, o RNA mensageiro recém-sintetizado pode ser imediatamente traduzido por ribossomos. Esse arranjo reduz o intervalo entre a ativação de um gene e a produção da proteína correspondente. Em ambientes instáveis, essa velocidade pode ser decisiva. Uma bactéria exposta a um novo substrato, a uma alteração de temperatura ou a um composto tóxico precisa ajustar rapidamente seu repertório proteico. A organização procarionte favorece esse tipo de resposta porque aproxima fisicamente informação genética e maquinaria biossintética [Figura].
O nucleoide também participa dessa integração de maneira ativa. O DNA circular compactado não é apenas um depósito de genes, mas uma estrutura dinâmica cujo grau de superenrolamento, posicionamento e associação com proteínas reguladoras influencia replicação, transcrição e divisão celular. A célula precisa coordenar a duplicação do cromossomo com o crescimento da membrana, a síntese da parede e a formação do septo. Se esses processos não estiverem sincronizados, a divisão pode gerar células inviáveis. Portanto, a reprodução procarionte depende da articulação entre arquitetura genética, crescimento físico e montagem estrutural [Quadro].
A divisão celular por fissão binária ilustra de forma clara essa lógica sistêmica. Antes da separação das células-filhas, o cromossomo precisa ser replicado, as cópias precisam ser posicionadas, o anel de FtsZ precisa se formar no local correto, a membrana precisa invaginar e a parede celular deve ser sintetizada de maneira coordenada. Cada etapa depende de sinais espaciais e moleculares que impedem divisão em locais inadequados ou antes da duplicação suficiente do material genético. A célula procarionte, portanto, não se divide apenas por “estrangulamento” simples; ela executa um processo ordenado de construção de duas novas unidades celulares [Figura].
As estruturas especializadas ampliam essa integração ao conectar a célula ao ambiente externo. Flagelos traduzem sinais químicos em movimento; fímbrias e pili transformam propriedades de superfície em adesão, colonização ou transferência genética; cápsulas e biofilmes convertem produção extracelular em proteção coletiva; endósporos reorganizam a célula para resistência extrema. Cada uma dessas estruturas depende de genes específicos, síntese proteica, gasto energético, montagem espacial e regulação ambiental. Por isso, motilidade, adesão, persistência e transferência genética não devem ser vistas como acessórios, mas como extensões funcionais da organização celular [Tabela].
A formação de biofilmes representa um caso particularmente expressivo de integração entre célula individual e comportamento coletivo. Uma bactéria aderida a uma superfície pode alterar sua expressão gênica, produzir matriz extracelular, comunicar-se quimicamente com outras células e estabelecer uma comunidade organizada. Nesse contexto, a função celular deixa de ser apenas individual e passa a depender da posição da célula dentro da comunidade, da disponibilidade local de oxigênio e nutrientes, da produção compartilhada de matriz e da tolerância coletiva a estresses. A célula procarionte revela, assim, que mesmo organismos unicelulares podem apresentar formas rudimentares de organização multicelular funcional [Figura].
O metabolismo é outro eixo de integração. As vias energéticas e biossintéticas precisam responder à disponibilidade de nutrientes, aceptores de elétrons, luminosidade, oxigênio e compostos químicos do ambiente. A célula ajusta a expressão de genes, a atividade de enzimas, a captação de substratos e a produção de resíduos conforme suas necessidades. Essa regulação evita desperdício energético e permite alternância entre diferentes modos metabólicos, como respiração, fermentação, fotossíntese ou quimiolitotrofia. A flexibilidade metabólica procarionte depende, portanto, da conexão entre sensores ambientais, regulação gênica, transporte de membrana e redes bioquímicas internas [Quadro].
Essa integração também explica a rapidez adaptativa dos procariontes. Populações bacterianas combinam divisão rápida, mutação, recombinação e transferência horizontal de genes. Plasmídeos, transposons, integrons e bacteriófagos podem introduzir novas capacidades funcionais, mas essas capacidades só se tornam biologicamente relevantes se forem integradas ao metabolismo, ao controle gênico e à estrutura celular. Um gene de resistência a antibiótico, por exemplo, pode codificar uma enzima inativadora, uma bomba de efluxo ou uma alteração no alvo molecular; em qualquer caso, seu efeito depende de expressão adequada, localização correta e compatibilidade com a fisiologia celular [Figura].
No ambiente, essa integração estrutural e funcional adquire escala ecológica. Bactérias do solo participam de ciclos de nutrientes porque sua organização celular permite captar substratos, processá-los metabolicamente e liberar produtos que serão utilizados por plantas ou outros microrganismos. Cianobactérias contribuem para produção primária porque integram pigmentos, membranas fotossintéticas, fixação de carbono e, em alguns casos, fixação de nitrogênio. Bactérias patogênicas causam doença porque coordenam adesão, invasão, evasão imune, produção de toxinas e resposta ao hospedeiro. Em todos esses casos, o impacto ecológico ou patológico nasce da integração de processos celulares básicos [Tabela].
A relação estrutura-função é, portanto, o princípio organizador da célula procarionte. A pequena dimensão favorece difusão rápida e elevada relação superfície-volume; a membrana concentra transporte e energia; a parede celular garante resistência e forma; o citoplasma denso acelera interações moleculares; o nucleoide acessível favorece resposta gênica rápida; os plasmídeos ampliam repertórios adaptativos; as estruturas de superfície conectam a célula ao ambiente; o metabolismo versátil permite explorar diferentes fontes de energia e matéria. Nenhum desses elementos explica sozinho o sucesso dos procariontes. O sucesso emerge da combinação entre todos eles [Figura].
Essa visão integrada também evita dois erros comuns no ensino de biologia celular. O primeiro é tratar a célula procarionte como uma célula incompleta, definida apenas pelo que não possui. O segundo é descrevê-la como uma lista de componentes sem mostrar como esses componentes trabalham juntos. Uma célula procarionte não é apenas “sem núcleo”, “sem mitocôndria” ou “sem organelas membranosas”. Ela é um sistema biológico completo, construído sobre uma lógica própria de compactação, velocidade, plasticidade genética e acoplamento funcional. Sua organização não é uma versão reduzida da célula eucarionte; é uma estratégia celular distinta [Quadro].
Compreender essa integração prepara o estudante para o próximo passo conceitual: comparar o modelo procarionte com o modelo eucarionte. Nas células eucariontes, muitos processos são separados por compartimentos membranosos, o que permite especialização, controle espacial refinado e aumento de complexidade interna. Nos procariontes, por outro lado, a eficiência depende da proximidade entre processos, da economia estrutural e da regulação rápida. A comparação entre esses dois modelos não deve ser feita em termos de superioridade, mas de estratégia organizacional. Ambos sustentam vida celular; cada um resolve os problemas de sobrevivência, reprodução e adaptação de maneira diferente.
Assim, a célula procarionte deve ser vista como um sistema integrado de alta eficiência. Sua organização estrutural favorece trocas rápidas com o ambiente, sua arquitetura genética permite respostas imediatas, seu metabolismo explora múltiplas fontes de energia, suas estruturas especializadas ampliam interação e sobrevivência, e sua plasticidade genética sustenta adaptação evolutiva. Esse conjunto explica por que bactérias e arqueias permanecem entre as formas de vida mais abundantes, antigas e bem-sucedidas do planeta. Mais que uma introdução à célula “simples”, o estudo da célula procarionte revela uma das soluções mais elegantes da biologia: fazer muito, em pouco espaço, com máxima integração funcional.
5.11 Comparação entre células procariontes e eucariontes #
A comparação entre células procariontes e eucariontes é um dos pontos centrais da biologia celular, porque permite compreender como diferentes modelos de organização podem sustentar a vida. Ambas compartilham propriedades fundamentais: são delimitadas por membrana plasmática, utilizam DNA como material genético, realizam síntese proteica por ribossomos, dependem de reações metabólicas para obtenção de energia e mantêm mecanismos de crescimento, reprodução e resposta ao ambiente. Essas semelhanças revelam uma base molecular comum, herdada de processos evolutivos antigos. Entretanto, a forma como essas funções são organizadas no espaço celular difere profundamente entre os dois tipos celulares, e é nessa diferença que se encontra a chave para compreender a transição da célula compacta procarionte para a célula compartimentalizada eucarionte [Figura].
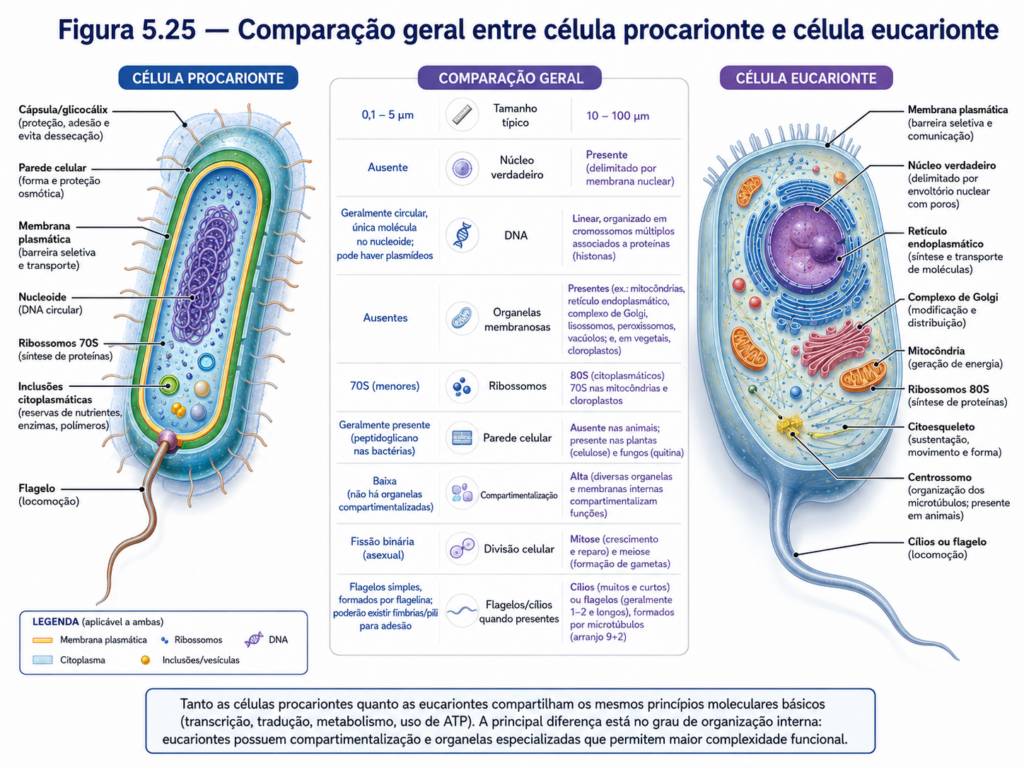
A distinção mais evidente está na organização do material genético. Nas células procariontes, o DNA encontra-se no nucleoide, uma região citoplasmática não delimitada por membrana, geralmente contendo um cromossomo circular associado a proteínas e organizado por superenrolamento. Nas células eucariontes, o DNA está contido no núcleo, envolvido por dupla membrana e organizado em cromossomos lineares associados a histonas, formando a cromatina. Essa diferença não é apenas morfológica. Ela modifica profundamente a dinâmica da expressão gênica. No procarionte, transcrição e tradução podem ocorrer de modo acoplado no citoplasma; no eucarionte, a transcrição ocorre no núcleo, o RNA é processado e exportado, e a tradução ocorre no citoplasma ou associada ao retículo endoplasmático rugoso [Figura].
Essa separação espacial nas células eucariontes permite níveis adicionais de controle. Antes que uma proteína seja sintetizada, o RNA mensageiro eucarionte pode sofrer capeamento, poliadenilação, remoção de íntrons, edição, exportação seletiva e regulação de estabilidade. Nos procariontes, a resposta tende a ser mais direta e rápida, com forte controle transcricional e tradução quase imediata. Portanto, a célula procarionte privilegia velocidade e economia; a célula eucarionte privilegia compartimentalização, processamento e regulação em múltiplas etapas. Nenhuma dessas estratégias é “melhor” em termos absolutos. Cada uma responde a pressões biológicas diferentes [Tabela].
Outra diferença fundamental está na compartimentalização interna. Células procariontes não possuem organelas membranosas clássicas, como mitocôndrias, cloroplastos, retículo endoplasmático, complexo de Golgi, lisossomos ou peroxissomos. Seus processos metabólicos ocorrem no citoplasma, na membrana plasmática ou em especializações membranosas. Já as células eucariontes distribuem funções em compartimentos internos especializados. A mitocôndria concentra processos de respiração celular; o cloroplasto realiza fotossíntese em células vegetais e algas; o retículo endoplasmático participa da síntese de proteínas e lipídios; o complexo de Golgi modifica e direciona moléculas; os lisossomos e peroxissomos executam funções degradativas e oxidativas específicas [Figura].
Essa compartimentalização eucarionte permite criar microambientes intracelulares com composição química, pH, concentração iônica, conjunto enzimático e função própria. Isso amplia o controle sobre reações incompatíveis entre si e permite maior especialização funcional. Em contraste, a célula procarionte opera com grande integração no mesmo espaço citoplasmático, dependendo de proximidade molecular, organização local e regulação rápida. O que no eucarionte é resolvido por separação física em compartimentos, no procarionte é resolvido por acoplamento funcional e organização molecular compacta [Quadro].
A diferença de escala também é relevante. Células procariontes costumam ser menores, frequentemente entre 1 e 10 micrômetros, embora existam exceções. Células eucariontes geralmente são maiores, muitas vezes entre 10 e 100 micrômetros. O tamanho reduzido dos procariontes favorece alta relação superfície-volume, difusão rápida e trocas eficientes com o ambiente. Nas células eucariontes, o aumento do volume exigiu soluções internas para transporte, organização e distribuição de moléculas. A compartimentalização e o desenvolvimento de um citoesqueleto complexo podem ser interpretados, em parte, como respostas a esse desafio físico: manter eficiência funcional em células maiores e internamente mais organizadas [Figura].
O citoesqueleto representa outro ponto de contraste. Durante muito tempo, acreditou-se que ele fosse uma característica exclusiva dos eucariontes. Hoje se reconhece que procariontes possuem proteínas estruturais relacionadas à manutenção da forma, divisão e organização interna, como FtsZ, MreB e outras. No entanto, o citoesqueleto eucarionte alcança maior complexidade, sendo composto por microtúbulos, microfilamentos de actina e filamentos intermediários, associados a proteínas motoras e a sistemas de transporte intracelular. Essa rede sustenta a forma celular, posiciona organelas, organiza a divisão celular, permite movimentos celulares e viabiliza transporte direcionado de vesículas e moléculas [Figura].
Na divisão celular, as diferenças também são marcantes. Procariontes dividem-se predominantemente por fissão binária, processo no qual ocorre replicação do cromossomo, segregação das cópias e formação de septo. Eucariontes realizam mitose para divisão de células somáticas e meiose para formação de gametas ou esporos em ciclos sexuados. A mitose exige condensação cromossômica, formação do fuso mitótico, ligação dos cromossomos aos microtúbulos, separação precisa das cromátides e citocinese. A meiose adiciona ainda recombinação genética e redução do número cromossômico. Essas diferenças refletem a maior complexidade do genoma e da organização cromossômica eucarionte [Tabela].
Os ribossomos oferecem uma comparação particularmente didática. Procariontes possuem ribossomos 70S, formados por subunidades 30S e 50S. Eucariontes possuem ribossomos citoplasmáticos 80S, formados por subunidades 40S e 60S. Essa diferença estrutural tem grande importância biomédica, pois muitos antibióticos atuam seletivamente sobre ribossomos bacterianos, interferindo na síntese proteica sem bloquear diretamente os ribossomos citoplasmáticos eucariontes. A presença de ribossomos semelhantes aos bacterianos em mitocôndrias e cloroplastos também reforça a hipótese endossimbiótica, segundo a qual essas organelas derivam de antigos procariontes incorporados por células ancestrais [Quadro].
A hipótese endossimbiótica é um dos principais pontos de ligação entre os dois modelos celulares. Segundo essa interpretação, mitocôndrias e cloroplastos teriam se originado de bactérias que passaram a viver no interior de uma célula ancestral eucariótica. Com o tempo, essa relação tornou-se obrigatória, com transferência de parte dos genes para o núcleo e integração funcional ao metabolismo celular. Evidências como dupla membrana, DNA próprio, divisão por processos semelhantes à fissão, ribossomos próprios e semelhanças bioquímicas com bactérias sustentam essa interpretação. Assim, a célula eucarionte não surge simplesmente como oposição ao procarionte, mas como resultado de eventos evolutivos que envolveram integração entre linhagens celulares distintas [Figura].
Também há diferenças importantes na parede celular. Muitas bactérias possuem parede rica em peptidoglicano, componente ausente nas células eucariontes. Arqueias podem apresentar paredes com composições distintas, sem peptidoglicano típico bacteriano. Entre os eucariontes, células vegetais possuem parede rica em celulose, fungos apresentam parede com quitina, enquanto células animais não possuem parede celular, dependendo de matriz extracelular, citoesqueleto cortical e junções celulares para suporte e organização tecidual. Essa comparação mostra que a presença de parede celular não é exclusiva dos procariontes, mas sua composição e função variam entre os diferentes grupos [Tabela].
A organização metabólica também segue lógicas distintas. Procariontes apresentam grande diversidade energética, incluindo respiração aeróbia, respiração anaeróbia, fermentação, fotossíntese oxigênica, fotossíntese anoxigênica, quimiolitotrofia e fixação de nitrogênio. Muitos desses processos ocorrem diretamente associados à membrana plasmática. Eucariontes, por sua vez, concentram a respiração aeróbia nas mitocôndrias e a fotossíntese nos cloroplastos, quando presentes. Embora eucariontes sejam estruturalmente mais compartimentalizados, a amplitude metabólica global dos procariontes é extraordinariamente maior. Esse é um ponto didático decisivo: complexidade estrutural e diversidade metabólica não são a mesma coisa [Quadro].
Do ponto de vista genômico, os procariontes tendem a apresentar genomas mais compactos, com alta densidade gênica, poucos íntrons e frequente organização de genes em operons. Os eucariontes, especialmente multicelulares, possuem genomas maiores, com maior proporção de sequências não codificadoras, íntrons, elementos repetitivos e regiões regulatórias extensas. Essa arquitetura permite regulação gênica complexa, diferenciação celular, desenvolvimento multicelular e controle espacial e temporal da expressão gênica. Nos procariontes, a organização genética favorece respostas rápidas e economia de recursos; nos eucariontes, favorece controle refinado e diversificação funcional entre tipos celulares [Figura].
A relação com o ambiente também difere. Células procariontes, especialmente organismos unicelulares livres, estão frequentemente em contato direto com variações ambientais e precisam responder rapidamente a mudanças de temperatura, pH, osmolaridade, nutrientes, toxinas e competição. Células eucariontes unicelulares também enfrentam esse tipo de desafio, mas células eucariontes de organismos multicelulares vivem frequentemente em ambientes internos regulados, integradas a tecidos, sistemas de comunicação e matriz extracelular. Assim, a célula eucarionte multicelular não é apenas uma célula mais complexa; ela é uma unidade funcional inserida em um organismo, sujeita a controles sistêmicos [Quadro].
Essa diferença ajuda a compreender a evolução da multicelularidade. Embora existam biofilmes e comportamentos coletivos em procariontes, a multicelularidade eucarionte alcançou níveis elevados de diferenciação, especialização tecidual e desenvolvimento embrionário. Isso se relaciona à compartimentalização, ao citoesqueleto complexo, à comunicação celular, à adesão intercelular, à regulação gênica refinada e à capacidade de gerar tipos celulares distintos a partir de um mesmo genoma. Procariontes formam comunidades altamente organizadas, mas não tecidos verdadeiros nos moldes de plantas e animais. Essa comparação deve ser feita com precisão para não reduzir biofilmes a “tecidos bacterianos” nem ignorar sua organização coletiva [Figura].
Apesar das diferenças, há uma profunda continuidade molecular entre os dois modelos celulares. O código genético é praticamente universal; ATP atua como moeda energética; DNA, RNA e proteínas ocupam posições centrais; ribossomos realizam tradução; membranas lipídicas delimitam o espaço celular; enzimas catalisam reações; gradientes eletroquímicos sustentam trabalho biológico. Essa unidade mostra que a vida celular compartilha princípios fundamentais, mesmo quando a arquitetura diverge. O estudo comparativo, portanto, não deve produzir uma oposição simplista entre “simples” e “complexo”, mas revelar como diferentes soluções celulares emergiram a partir de uma base comum [Tabela].
Em termos didáticos, a comparação entre procariontes e eucariontes cumpre uma função estratégica no percurso do livro. Até aqui, a célula procarionte foi apresentada como um sistema compacto, rápido, metabolicamente diverso e geneticamente plástico. No próximo capítulo, a célula eucarionte será examinada como um sistema compartimentalizado, internamente especializado e estruturalmente mais elaborado. A transição entre esses capítulos deve ser entendida como mudança de modelo organizacional: da integração direta em pequeno volume para a distribuição de funções em compartimentos internos [Figura].
Portanto, células procariontes e eucariontes representam duas grandes estratégias de organização da vida celular. Os procariontes mostram como uma célula pequena, sem núcleo e sem organelas membranosas clássicas pode alcançar enorme eficiência ecológica, metabólica e evolutiva. Os eucariontes mostram como a compartimentalização interna, o núcleo, o citoesqueleto complexo e o controle gênico refinado permitem aumento de tamanho, especialização e multicelularidade complexa. A comparação entre esses dois modelos não diminui nenhum deles; ao contrário, evidencia a diversidade de caminhos pelos quais a vida resolveu os mesmos problemas fundamentais: proteger a informação genética, obter energia, produzir moléculas, responder ao ambiente, reproduzir-se e persistir ao longo do tempo.
5.12 Perspectivas modernas em biologia procarionte #
O estudo dos procariontes deixou de ser restrito à descrição de bactérias cultivadas em laboratório ou à comparação clássica entre células com e sem núcleo. Nas últimas décadas, a biologia procarionte passou a ocupar uma posição central em áreas como microbioma, ecologia molecular, resistência antimicrobiana, biologia sintética, agricultura sustentável, biorremediação e astrobiologia. Essa mudança ocorreu porque novas tecnologias permitiram observar não apenas células isoladas, mas comunidades microbianas inteiras, seus genes, suas interações metabólicas e sua influência sobre organismos, ambientes e sistemas produtivos. A célula procarionte, antes apresentada muitas vezes como modelo simples de organização celular, tornou-se uma das chaves para compreender processos biológicos em escala planetária [Figura].
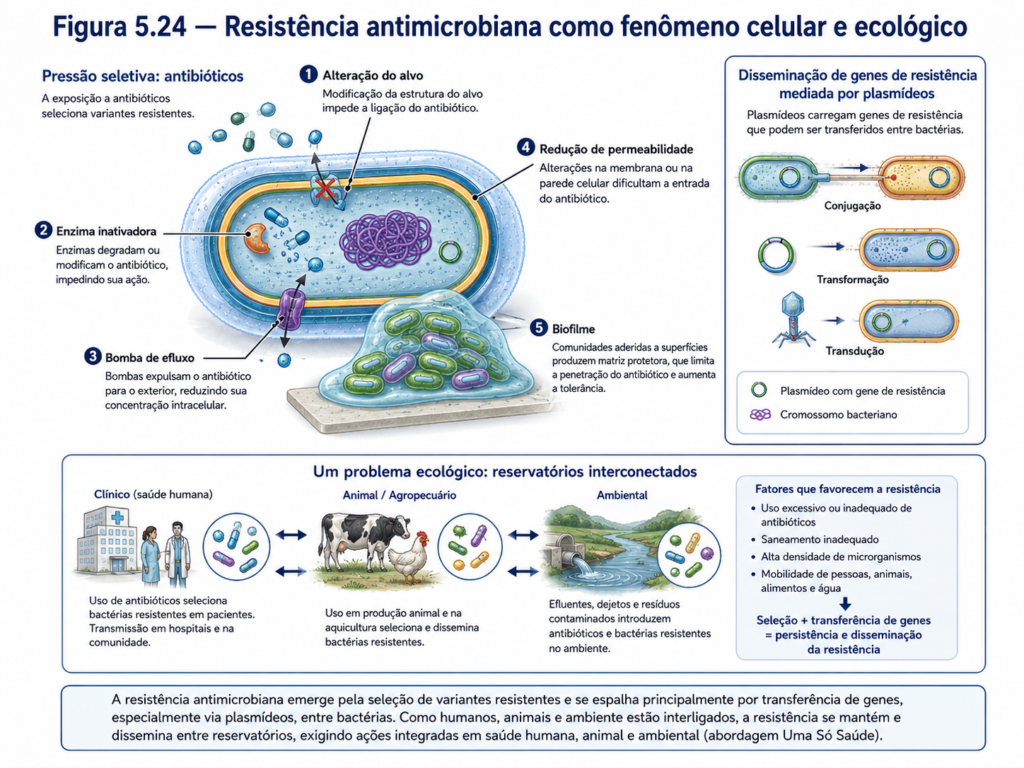
Uma das principais transformações modernas foi a expansão do conceito de microbioma. O termo não se refere apenas à presença de microrganismos em determinado ambiente, mas ao conjunto de comunidades microbianas, seus genomas, seus produtos metabólicos e suas interações com o hospedeiro ou com o ecossistema. Em seres humanos, animais, plantas, solos, águas, alimentos e ambientes construídos, bactérias e arqueias formam redes complexas que modulam processos fisiológicos, ecológicos e produtivos. Assim, a biologia celular procarionte passou a ser estudada não apenas no nível da célula individual, mas também no nível das comunidades e de suas funções coletivas [Figura].
No caso das plantas, o microbioma associado à rizosfera, à filosfera, às sementes e aos tecidos internos influencia crescimento, nutrição, tolerância a estresses e resistência a patógenos. Bactérias promotoras de crescimento vegetal podem produzir fitormônios, solubilizar nutrientes, fixar nitrogênio, modular respostas imunes ou competir com microrganismos prejudiciais. Em ambientes semiáridos, onde a limitação hídrica, a salinidade e a baixa matéria orgânica frequentemente restringem a produtividade, compreender essas interações pode abrir caminhos para bioinsumos mais eficientes e manejos mais adaptados à realidade local. Nesse cenário, o estudo dos procariontes conecta biologia celular, agronomia, ecologia do solo e inovação tecnológica [Quadro].
A metagenômica foi decisiva para essa mudança de perspectiva. Durante muito tempo, a microbiologia dependia do cultivo de microrganismos em meios artificiais, o que limitava fortemente a visão da diversidade procarionte, já que grande parte das bactérias e arqueias ambientais não cresce facilmente em condições laboratoriais convencionais. Com o sequenciamento direto de DNA extraído de amostras ambientais, tornou-se possível investigar comunidades inteiras sem necessidade de cultivo prévio. Essa abordagem revelou uma diversidade microbiana muito mais ampla, além de genes, vias metabólicas e relações ecológicas que permaneciam invisíveis aos métodos tradicionais [Figura].
Além da metagenômica, outras abordagens ômicas ampliaram a compreensão dos procariontes. A metatranscriptômica permite avaliar quais genes estão sendo expressos em uma comunidade; a metaproteômica identifica proteínas produzidas em determinado contexto; a metabolômica descreve compostos gerados ou consumidos por microrganismos; e a genômica de célula única permite estudar organismos ainda não cultivados. Integradas, essas ferramentas ajudam a sair de uma pergunta descritiva — “quem está presente?” — para perguntas funcionais mais relevantes: “o que esses microrganismos estão fazendo?”, “como interagem?”, “como respondem ao ambiente?” e “como podem ser manejados?” [Tabela].
Outra perspectiva moderna relevante é a compreensão da resistência antimicrobiana como fenômeno ecológico, evolutivo e celular. A resistência não surge apenas em hospitais ou laboratórios; genes de resistência circulam naturalmente em solos, águas, microbiomas animais, ambientes agrícolas e sistemas de tratamento de resíduos. O uso intensivo de antimicrobianos em saúde humana, veterinária e produção agropecuária aumenta a pressão seletiva sobre populações bacterianas, favorecendo variantes capazes de sobreviver. Como muitos genes de resistência podem estar em plasmídeos, transposons, integrons e outros elementos móveis, a disseminação não depende apenas da reprodução clonal, mas também da transferência horizontal entre microrganismos [Figura].
Essa visão desloca o problema da resistência antimicrobiana para uma abordagem integrada. Não basta pensar no antibiótico e na bactéria isoladamente. É necessário considerar o ambiente, o hospedeiro, o uso de medicamentos, o manejo agropecuário, a qualidade da água, o destino de resíduos, a densidade microbiana, os biofilmes e as rotas de transferência genética. A célula procarionte, com sua parede, membrana, bombas de efluxo, ribossomos, plasmídeos e mecanismos regulatórios, está no centro desse processo. Porém, o fenômeno só se explica plenamente quando esses componentes celulares são analisados dentro de populações e ecossistemas [Quadro].
No campo da biologia sintética, os procariontes passaram a ser tratados como plataformas celulares programáveis. Bactérias podem receber circuitos genéticos artificiais, sensores moleculares, vias metabólicas reconstruídas e sistemas regulatórios projetados para executar funções específicas. Essa abordagem permite imaginar células capazes de detectar contaminantes, produzir moléculas de interesse, responder a sinais ambientais, atuar como biossensores, fabricar biopolímeros, converter resíduos em produtos úteis ou auxiliar no diagnóstico de doenças. O princípio é simples em sua formulação, mas tecnicamente complexo: utilizar a maquinaria celular procarionte como base para sistemas biológicos redesenhados [Figura].
A engenharia metabólica é uma das áreas mais diretamente associadas a essa perspectiva. Ao modificar genes, promotores, enzimas e fluxos metabólicos, é possível redirecionar a célula bacteriana para produzir compostos de interesse industrial, farmacêutico, agrícola ou ambiental. A eficiência desse processo depende de uma compreensão fina da fisiologia celular: a célula precisa crescer, manter estabilidade genética, expressar as enzimas desejadas, evitar acúmulo de intermediários tóxicos, equilibrar consumo de energia e conservar viabilidade. Esse ponto é didaticamente importante porque mostra que uma bactéria não é um “recipiente” passivo para genes; ela é um sistema vivo com restrições, prioridades metabólicas e respostas regulatórias próprias [Quadro].
As tecnologias baseadas em CRISPR também modificaram a biologia procarionte. Originalmente, sistemas CRISPR-Cas foram identificados como mecanismos de defesa de bactérias e arqueias contra elementos genéticos invasores, como vírus e plasmídeos. A partir da compreensão desse sistema natural, pesquisadores desenvolveram ferramentas de edição genômica amplamente utilizadas em diferentes organismos. Esse caso ilustra de maneira exemplar como o estudo de um mecanismo celular procarionte pode gerar uma tecnologia transformadora para biologia molecular, medicina, agricultura e biotecnologia [Figura].
A relação entre procariontes e vírus bacterianos, especialmente bacteriófagos, também voltou ao centro da pesquisa contemporânea. Fagos influenciam a composição de comunidades bacterianas, transferem genes, regulam populações microbianas e podem ser explorados como alternativas ou complementos ao controle de bactérias patogênicas. A fagoterapia, embora antiga em sua origem, tem recebido atenção renovada diante da resistência antimicrobiana. Seu uso exige precisão, pois a especificidade entre fago e bactéria, a evolução de resistência bacteriana aos fagos, a resposta imune do hospedeiro e a segurança do tratamento precisam ser cuidadosamente avaliadas [Quadro].
Na agricultura, o interesse moderno por procariontes também está ligado ao desenvolvimento de bioinsumos. Inoculantes, biofertilizantes, solubilizadores de nutrientes, agentes de biocontrole e consórcios microbianos são cada vez mais discutidos como alternativas ou complementos aos insumos químicos convencionais. Contudo, a eficácia desses produtos depende de validação experimental rigorosa. É necessário demonstrar que a cepa ou o consórcio permanece viável, coloniza o ambiente-alvo, desempenha a função desejada, interage adequadamente com a cultura e mantém desempenho sob condições reais de campo. Sem isso, o discurso biotecnológico pode se transformar em promessa agronômica frágil [Tabela].
Essa observação é especialmente relevante porque o mercado de produtos microbiológicos cresce rapidamente, mas nem toda formulação tem a mesma qualidade. A biologia celular fornece critérios para análise: o microrganismo precisa resistir ao armazenamento, suportar aplicação, sobreviver ao ambiente, competir ou cooperar com a microbiota local e expressar os mecanismos funcionais esperados. Um produto que contém células inviáveis, cepas mal caracterizadas ou concentrações inadequadas dificilmente entregará resultado consistente. Portanto, compreender os procariontes em nível celular protege o futuro profissional contra interpretações superficiais e decisões técnicas baseadas apenas em apelo comercial [Quadro].
Outra fronteira moderna envolve o estudo de procariontes em ambientes extremos e sua relação com a astrobiologia. A presença de bactérias e arqueias em fontes hidrotermais, desertos, geleiras, lagos hipersalinos, ambientes ácidos e regiões profundas amplia a compreensão sobre os limites físico-químicos da vida. Essa informação é útil para investigar a origem da vida na Terra e para formular hipóteses sobre possíveis formas de vida em outros corpos planetários. Extremófilos mostram que células podem manter organização molecular, metabolismo e reprodução sob condições muito diferentes daquelas consideradas ideais para organismos multicelulares complexos [Figura].
A biotecnologia industrial também se beneficia dessa diversidade extrema. Enzimas termoestáveis, proteínas resistentes a salinidade, sistemas metabólicos anaeróbios e moléculas adaptadas a condições severas podem ser explorados em processos industriais, diagnósticos, produção de biocombustíveis, tratamento de resíduos e síntese química. A utilização de DNA polimerases termoestáveis em técnicas de amplificação de DNA é um exemplo clássico de como uma adaptação celular de microrganismos termófilos pode sustentar uma tecnologia laboratorial de enorme impacto [Quadro].
A perspectiva moderna também exige reconhecer que procariontes não vivem como unidades isoladas na maior parte dos ambientes naturais. Eles formam consórcios, biofilmes, redes metabólicas e associações simbióticas. Em um biofilme, células podem apresentar diferentes estados fisiológicos de acordo com sua posição, acesso a oxigênio, nutrientes e sinais químicos. Em comunidades do solo, uma espécie pode liberar metabólitos que alimentam outra, enquanto uma terceira modifica o pH, o oxigênio ou a disponibilidade de minerais. Esse comportamento coletivo obriga a biologia celular a dialogar com ecologia, modelagem de sistemas e análise de redes [Figura].
Essa mudança tem implicações para o ensino. O estudante precisa conhecer a estrutura celular básica, mas também precisa entender que a célula procarionte é parte de populações e comunidades. Uma bactéria em placa de Petri não representa, sozinha, a complexidade de uma comunidade rizosférica, de um biofilme em tubulação de irrigação, de um microbioma intestinal ou de um sedimento anaeróbio. A célula individual continua sendo a unidade fundamental, mas seu comportamento real depende frequentemente do contexto coletivo. Essa tensão entre célula e comunidade é uma das marcas da biologia procarionte contemporânea [Quadro].
As perspectivas modernas também reforçam a necessidade de integrar ciência básica e aplicação. Não há bioinsumo confiável sem microbiologia celular robusta; não há enfrentamento sério da resistência antimicrobiana sem compreender genética bacteriana; não há biologia sintética segura sem fisiologia celular; não há manejo sustentável do solo sem ecologia microbiana; não há uso responsável de organismos modificados sem biossegurança. A célula procarionte, portanto, funciona como ponto de convergência entre pesquisa fundamental, tecnologia e tomada de decisão em sistemas reais [Tabela].
Nesse contexto, o professor e o estudante precisam evitar duas simplificações. A primeira é tratar bactérias apenas como agentes de doença. A segunda é tratá-las apenas como ferramentas úteis. Procariontes são organismos com história evolutiva própria, capazes de interagir, adaptar-se, persistir e transformar ambientes. Podem sustentar ciclos de nutrientes, proteger plantas, produzir medicamentos, degradar resíduos e compor microbiomas saudáveis; mas também podem causar doenças, disseminar resistência, contaminar processos produtivos e gerar desequilíbrios ambientais. A leitura madura da biologia procarionte exige manter essas dimensões simultaneamente [Quadro].
Assim, as perspectivas modernas em biologia procarionte ampliam o alcance deste capítulo. O que começou com a descrição de uma célula pequena, sem núcleo delimitado por membrana, termina como uma visão integrada de organismos que sustentam ciclos ecológicos, influenciam saúde, alimentam tecnologias e desafiam modelos tradicionais de organização biológica. Bactérias e arqueias mostram que a vida pode ser compacta, rápida, cooperativa, resistente, editável, comunitária e profundamente transformadora. Compreender essa lógica prepara o estudante para avançar ao estudo da célula eucarionte sem enxergá-la como simples “evolução superior”, mas como outra estratégia celular, construída sobre compartimentalização, especialização e integração interna.
Estudo dirigido — Capítulo 5: Célula Procarionte #
- Explique por que a célula procarionte não deve ser considerada uma célula “simples” apenas por não possuir núcleo delimitado por membrana nem organelas membranosas clássicas.
- Descreva as principais diferenças estruturais entre bactérias Gram-positivas e Gram-negativas, relacionando essas diferenças com permeabilidade, proteção celular e sensibilidade a antibióticos.
- Analise como a ausência de núcleo nos procariontes permite o acoplamento entre transcrição e tradução, e explique a vantagem funcional desse processo para a resposta rápida ao ambiente.
- Compare fissão binária, conjugação, transformação e transdução, destacando quais processos estão relacionados à reprodução celular e quais estão relacionados à variabilidade genética.
- Explique como a diversidade metabólica dos procariontes contribui para sua presença em diferentes ambientes e relacione esse fato à agricultura, especialmente ao ciclo do nitrogênio e à fertilidade do solo.
Questão para estimular pesquisa #
- Pesquise um exemplo atual de aplicação de procariontes na agricultura, saúde ou biotecnologia — como bioinsumos, biorremediação, produção de proteínas recombinantes, CRISPR-Cas ou resistência antimicrobiana — e explique quais características celulares procariontes tornam essa aplicação possível.



